МЕХАНИЗМЫ ВЫЖИВАНИЯ И РАСПРОСТРАНЕНИЯ ПАТОГЕННЫХ МИКРООРГАНИЗМОВ (ГИПОТЕЗА)
В. Е. Жемчугов, Межрегиональный институт иммунокоррекции и метаболической терапии, Москва Взаимодействие микробов и человека находится в центре внимания ученых (врачей, биологов и философов) со времени открытия микробов. Как биолог-теоретик, так и практический врач должны знать, в какой степени закономерности развития популяции микробов в пробирке, ферментере, на чашке Петри (in vitro) соответствуют процессу эволюции микроорганизмов в природе (in vivo), а также в организме хозяина в процессе заражения. Существует ли единая “стратегия” развития микробного типа, популяции?Целью настоящего исследования является изучение некоторых закономерностей популяционного типа микробного сообщества в зависимости от фаз роста, чтобы определить, в какой момент жизненного цикла микробной популяции она может быть затронута в интересах человека. Для иллюстрации остановимся на возбудителях чумы, туляремии, мелиоидоза и сап, которые были в центре моих научных исследований в течение 20 лет. В отличие от микробов, существовавших на Земле миллиарды лет назад, человек как вид живет всего миллионы лет. Поэтому неуместно ставить homo sapiens в центр любых изменений или прогресса популяции микроорганизмов.
Классическая эпидемиология рассматривает эпидемический или эпизоотический (среди животных) процесс как цепочку, состоящую из источника, пути передачи, восприимчивого организма, который часто после заражения оказывается одновременно источником – триада Громашевского /17/.
Следует отметить, что в зависимости от экологии источника можно выделить антропонозы, зоонозы и сапронозы /8/. В свою очередь, микроорганизмы делятся на облигатно - патогенные, условно-патогенные и сапрофитные /9/.
Согласно большинству эпидемиологических классификаций, человек рассматривается как основной объект для развития инфекционного процесса с участием определенного типа микроорганизмов. Эпизоотия чумы трактуется аналогичным образом, как непрерывная цепь заболеваний восприимчивых животных, распространяющих микробы друг на друга любым способом.
С дальнейшим прогрессом человеческой инфекции или параллельно с первой, в человеческой популяции, этот процесс будет развиваться и уже будет называться эпидемическим.
Для Francisella tularensis человек обычно считается тупиковым, поскольку распространение от человека к человеку, в принципе невозможно. Популяция микроба мелиоидоза развивается без особых различий как in vivo, так и на белковых субстратах, например, растворенные в воде рисовые поля, болота. Кроме того, не происходит адаптации микроба мелиоидоза к какому-либо конкретному хозяину, поэтому процесс его взаимодействия с макроорганизмом можно рассматривать как сапроноз /7/.
Если мы примем во внимание, что все указанные типы микроорганизмов существовали задолго до появления человека на Земле, паразитируя на животных, которые уже сформировались как виды, необходимо будет признать, что популяция микробов на указанной территории является единым целым. Сущность объединяет общая эволюционная тенденция, “стратегия”, в основном “игнорирующая” близкое присутствие человеческого сообщества. С учетом этого, на наш взгляд, целесообразно рассматривать их взаимодействие, исходя из “микробоцентрических” позиций, а не из “антропоцентрических”.
Общий закон биологии, “стратегия” вида, направлена на его сохранение / выживание и распространение, то есть захват новых реалов. По нашему мнению, этот закон в полной мере можно отнести к популяции и видам микробов в целом. С этой точки зрения мы остановимся на процессах распространения некоторых типичных патогенных микробов.
Возбудители чумы, туляремии, сап и мелиоидоза, четко реализуют двухэтапную “стратегию” выживания и экспансии вида.
Первый шаг - это рост численности популяции в конкретной среде: либо это отдельный индивид (человек или животное), либо царство в целом.
Второй шаг - это прыжок в новое царство. Это может быть передача другому организму тем или иным способом или переход на другую территорию с помощью организма - носителя, природных или техногенных факторов, например, спор микробов сибирской язвы в перемещенной почве.
Могут ли микроорганизмы оказывать глубокое влияние на условия их распространения? Проникновение микробов в макроорганизм, скорее всего, будет пассивным вероятностным процессом, определяемым влиянием любых факторов, приводящих к переходу некоторых микробов в новое царство. Их судьба на новой территории будет зависеть от количества и “качества” клеток живого микроба в инокуляте, а также от условий in vivo (рис. 1)
С точки зрения популяции, “практичности” видов, т. е. полезности для видов в целом, “стратегия” культуры микробов может иметь два направления.
Первый: сохранение и расширение существующего царства,
второй : вторжение в новые царства. В первом направлении популяция должна достичь максимально возможной плотности клеток в условиях конкретной области. Фактически первый этап развития населения - это подготовка ко второму этапу. И потенциал, который будет приобретен на первом этапе, определит результат второго этапа: расстояние до нового царства, которое будет заселено. Кроме того, гибкость вида, потенциал его приспособляемости к различным условиям нового царства определяется генетической неоднородностью популяции, гетерогенностью, аналогичной генетике человека –его генотипом и “нормой реакции”. Второй этап – вторжение в новое царство – может быть реализован двумя способами. Первый способ, пассивный, предполагает превращение микроба в форму, удобную для сохранения в неблагоприятных условиях окружающей среды, и переход в новое царство, например, в споры. Часть микроорганизмов при достижении порога плотности для данной области, начинают синтезировать бактериоцины в качестве вторичных метаболитов, к которым также относятся многие антибиотики, продуцируемые в основном грибами. С точки зрения “популяционной целесообразности ” функция вторичных метаболитов заключается в защите границ царств от конкурирующих видов, а иногда, в ограничении размножения самой популяции (некоторых бактериоцинов), ее переключении на реализацию программы второго этапа – перехода в новое царство.
Таким образом, в отличие от зоонозов, существующих в природе независимо от наличия человека в их царстве и, эволюция которых коррелирует с “микробоцентрической” моделью, инфекции антропоноза, например, природная оспа, полностью описываются антропоцентрической моделью. Оба реализуют вышеупомянутую двухэтапную “стратегию” выживания и распространения в природе. Нет никаких сомнений в том, что развитие популяции зависит от видовых особенностей микробов, таких как фактор вирулентности, скорость размножения, генетическая гетерогенность и т. д.
В то же время, по мере увеличения численности населения, т. е. фаза его роста меняется, сами популяции создают условия для реализации второго этапа “стратегии” вида – перехода в другую сферу.
В контексте возбудителей чумы, туляремии, сапа и мелиоидоза представляет интерес изучение механизма реализации “межфазного” периода, в течение которого, популяция микробов не размножается и не погибает.
Если условия не позволяют размножаться вегетативным формам, а у микробов нет споровых форм, как они могут выживать в неблагоприятных для вегетативной формы условиях?
За последние годы, с помощью ДНК - РНК - диагностики (ПЦР), были выявлены “некультивируемые формы” для многих патогенных бактерий. Природа их сущности, механизмы формирования, существования, возврата к состоянию “культивирования” / сохранения до сих пор неизвестны.
Можно предположить, что такая нестабильная структура, как нуклеиновая кислота, может сохраняться только путем непрерывного размножения, в противном случае её гидролиз неизбежен. Хотя бесклеточные системы, воспроизводящие нуклеиновые кислоты, известны, трудно представить механизм их функционирования в природе. Вирусологи из Сергиево - Пасадского центра приложили огромные усилия для изучения возможностей реактивации вирусов оспы, обнаруженных в останках людей, умерших от этой болезни сотни и тысячи лет назад. К счастью, такие возможности не были раскрыты /14/. Поэтому сохранение наследственной субстанции патогенных типов в составе генома других видов, то есть эволюционных предшественников, приспособившихся к существованию в естественных условиях без паразитизма, представляется наиболее реалистичной гипотезой. Предположительный список таких пар для множества видов может быть составлен. Например, plage – иерсиниоз, холерный вибрион – НАГ - вибрион, возбудитель сибирской язвы – антракоиды, дифтерия – дифтероиды или коринебактерии, F. tularensi – непатогенная иерсиния, возможно, пастерелла, сапа - мелиоидоз – почвенные псевдомонады, возбудитель дизентерии – E. кишечная палочка и т. д.
Во всех случаях, можно предположить гипотетическую сборку исходного патогенного генома на основе непатогенного; рост / размножение и распространение в пределах области (индивидуума); прыжок в другую область, когда это невозможно - переход в “сохраненное” состояние, отдельное существование патогенных и непатогенных частей генома патогенного микроба.
При эпидемиологическом анализе участка, например, холеры, это выглядит как измененное соотношение изолированных вирулентных штаммов и, так называемых, НАГ - вибрионов. В дополнение к “непатогенной” и “патогенной” частям, может существовать третья запускающая часть генома или комбинация этой функции с любой из первых двух. В настоящее время, на наш взгляд, единственным подтверждением этой гипотезы для патогенных микроорганизмов, является возврат некультурной формы в обычное “культивируемое” состояние только путем прохождения через организм модельного животного, где, вероятно, произойдет “сборка” генома патогенного микроба. Более того, его наиболее легкая реализация происходит у высокочувствительных животных I группы, что касается восприимчивости и чувствительности, в соответствии с терминологией, принятой для туляремии /12, 20/. Популяция подобных животных является “легковоспламеняющимся материалом” для накопления микробов, а менее восприимчивые животные предназначены для их распространения на другие территории. Такие пары животных могут быть выбраны практически для всех зоонозных или антропонозных инфекций.
Для иллюстрации, остановимся на анализе динамики развития популяции возбудителей туляремии, чумы и мелиоидоза. В качестве ориентира, примем количество бактерий во внешней среде, достаточное для заражения восприимчивых животных I группы, что соответствует латентной фазе на кривой роста периодической культуры in vivo (см. рис. 1).
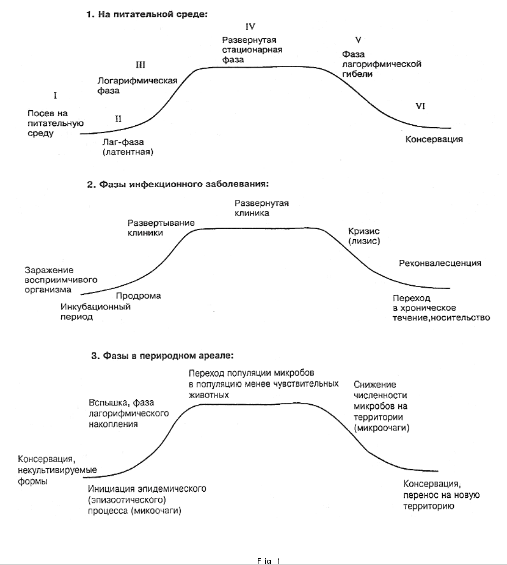
При минимальном количестве бактерий во внешней среде или минимальном количестве высокочувствительных и чувствительных животных на определенном пространстве единичные эпизоды инфекции происходят одновременно на всей территории микробного царства или участка.Согласно терминологии эпидемиологов по борьбе с чумой, это “небольшие участки” или микросайты. Когда популяция микробов достигает определенной концентрации или/и количество восприимчивых животных увеличивается, на территории начинается логарифмическая фаза,общее количество микробов увеличивается как у животных, так и во внешней среде. Согласно терминологии, используемой в настоящей статье, это реализация первого этапа демографической “стратегии”. Когда микробы достигают определенной плотности на территории, задействуются горизонтальные пути передачи, в том числе с использованием носителей, и становится возможным и вероятным, наряду с прогрессированием заболевания, заражать менее восприимчивых и чувствительных животных II группы (в отношении туляремии). Напомним, что человек также принадлежит к той же группе. Животные с меньшей чувствительностью к инфекции не умирают после заражения, но могут перемещаться на любое расстояние и каким - то образом оставлять там во внешней среде микробы, которые способны либо сохраниться, либо размножиться и начать вторгаться в новое царство. Развитие ситуации зависит от тех же причин: количество микробов , вторгшихся на новую территорию, “инфекционная доза”, их состояние (фаза развития), статус территории (“восприимчивая” или “иммунная”). Развитие популяции микробов In vivo также зависит от дозы заражения, состояния микроба, состояния макроорганизма, воздействия внешней среды, а также от профилактического и терапевтического воздействия как на микро -, так и на макроорганизм.
Скрытая фаза преобразуется либо в фазу логарифмического умножения, либо приводит к уничтожению микроба. Хотя иногда латентная фаза длится довольно долго (мелиоидоз), а для начала логарифмического размножения требуется серьезный сбой в функционировании макроорганизма.
Фазы клинического течения заболевания, как правило, хорошо совместимы с фазами развития популяции микробов: инкубационный период - латентная фаза, продромальный период - начало размножения, полномасштабная клиническая картина соответствует логарифмической фазе роста и стационарной фазе, кризис или лизис совместимы с началом деструкции и фазой логарифмической деструкции микроба.
Популяционная “стратегия” после достижения определенной численности, качественно меняется, часто, в ущерб “интересам” и жизни индивида, члена сообщества. Макроскопическим примером является переход пчёл или муравьёв, в состояние роения, после того, как пчелиная колония достигла критической численности в конкретном улье (муравейнике), уничтожение трутней. Существует множество, довольно ярких примеров реализации подобной “стратегии”, среди многочисленных типов микроорганизмов.
Анаэробы “выбрали” для себя пассивную позицию: в макроорганизме, доведенном до смерти мощными экзотоксинами, происходят изменения значений рН, температуры, содержания кислорода, а многочисленная популяция (колония) годами и веками переходит в споры (сибирская язва, ботулизм, газовая гангрена) /18/.
В то же время, имеются примеры активного влияния микроорганизмов на процесс их сохранения и распространения, с помощью неизвестных до сих пор механизмов. Вполне возможно, что популяция микробов адаптируется, чтобы соответствовать определенному механизму передачи /17/.
Врачи знают, что одним из симптомов бешенства, является интенсивное слюноотделение. В то же время, даже изначально неагрессивные животные, когда они заболевают, начинают кусать всё и всех, с кем сталкиваются. В сообщённых исследованиях болезни, врачи описали непреодолимое желание укусить, даже очень близкого человека. Что это? Каков механизм возникновения такого желания? Неоспоримым фактом является то, что именно в слюне содержится огромное количество вирусов бешенства.
Другой пример - грипп. Основной, носоглоточный, синдром (кашель, чихание), подразумевает, что окружающая среда вокруг больного человека заражена вирусами. Или холера: основным симптомом является сильнейшая диарея и рвота. Обе жидкости, на определенной стадии, представляют собой в основном, чистую высококонцентрированную культуру вибрионов. Используя терминологию Дарвина, возникает вопрос: кто к чему “приспособился"?
И, наконец, есть ключевой вопрос, непосредственно связанный с изучаемыми микробами – чумой, туляремией, эквинией и мелиоидозом : что произойдет, если “цель”, конечная цель второго этапа развития популяции микробов, не будет достигнута? Культура, накопленная внутри инфицированного высокочувствительного макроорганизма, не могла напрямую или через носителей проникнуть в макроорганизм, менее чувствительный, способный передать ее в новое царство или даже в аналогичный высокочувствительный организм, чтобы накопление микробов на данной территории продолжалось, и общий “легковоспламеняющийся материал сгорел”. После уничтожения животного, как жизнеспособные, так и нежизнеспособные микробы попадают во внешнюю среду.
Мы уже останавливались на спорообразователях, они реализуют “стратегию” пассивного сохранения и расширения. Но если споры не образуются? Многолетние споры о механизме выживания чумного микроба в межэпизоотический период привели в настоящее время к отсутствию каких-либо сообщений. Этот спокойный период можно объяснить попыткой понять роль “некультивированных” форм в эволюции не только чумного микроба, но и других микроорганизмов. Существование бактерий на объектах окружающей среды в форме неживой, но способной возвращаться к жизнеспособному, размножающемуся, вирулентному микробу, так называемыми "некультивированными” формами, можно считать доказанным фактом /1,9-13,15,16/. На самом деле, это то, что реагирует с зондом нуклеиновой кислоты в ПЦР или аналогичных системах и, не может расти на искусственных питательных средах.
Некоторые авторы сообщают о возвращении этого “чего-то” к первоначальной форме, посредством прохождения через организм модельного животного /12/. Никому не удалось воспроизвести in vitro весь процесс перехода в некультивированное состояние и обратно, с фиксацией переходных моментов. Поэтому ситуация выглядит как черный ящик. Видно, что перед ним и позади, но не известно, что внутри.
Другое понимание механизма выживания / сохранения чумного микроба в межэпизоотический период, было сообщено и интенсивно распространено, и разработано А. И. Дятловым /4,5/. Его гипотеза или теория, оправдывает процесс неоформления, в частности чумного микроба, не только в геологическую эпоху, но и в течение текущего периода. Не нужно спорить с тем фактом, что эволюционный процесс на планете, всё ещё продолжается, он ещё не закончился, когда появилась “вершина эволюции” - человек. И логично предположить, что если какой - либо микроб, давным - давно, в результате какого - либо процесса, в его генетическом аппарате, в определенных условиях окружающей среды, приобрел новые свойства, которые были развиты в следующих поколениях этого микроба, то все может произойти и происходит в настоящее время. Доступные статьи, посвященные открытию некультивированных форм /1,9-13,15/ не отвечают на вопрос, в какой форме существуют фрагменты нуклеиновых кислот, обнаруженные с помощью ПЦР. Разумно сделать вывод, что микроорганизмы находятся в некультивированной форме. Значительные фрагменты ДНК или м-РНК, переносящие РНК, очень маловероятно, чтобы автономно существовали, достаточно долго, во внешней среде. Как и чем их можно защитить? В литературе, нам не удалось найти ответов на эти вопросы. Имеются сообщения о существовании живых микробов (но не их нуклеиновых кислот) в растениях, простейших /7, 19/, водных организмах. Было установлено, что они не были “голыми”, т. е. клеточными нуклеиновыми кислотами, не содержащие мембран и, конечно, их фрагменты.
Существует еще один сообщенный факт о механизме мобилизации участков ДНК бактериальных клеток, с помощью транспозонов, а также обмена плазмидами микроорганизмов. Что касается вирусов, то давно, с помощью фагов, было сообщено, что их нуклеиновые кислоты обладают значительным потенциалом подвижности или инвазивности. Они способны не только вторгаться в прокариоты и эукариоты, и их геном, но и “тащить” за собой чьи - то (предыдущего хозяина) участки ДНК/РНК /16/.
Следовательно, со смертью хозяина, у патогенных микробов, есть следующие варианты выживания во внешней среде, помимо образования спор: они могут выжить, например, путем деления его генома при разрушении. Одна часть, вероятно, обладает способностью к сапрофитному существованию во внешней среде (Yersinia pseudotuberculosis) /8/, а вторая часть, определяющая способность паразитировать на живых организмах, сохраняется в виде мобилизованных транспозонами плазмид или фрагментов ДНК, встроенных в любой организм, бактерию, одноклеточный организм и т. д. Предположив существование такого механизма, мы увидим, что цепь замкнута со всеми доступными звеньями (рис. 2).
Таким образом, для начала реализации первого этапа гипотетического цикла развития популяции патогенных микроорганизмов, необходимы следующие компоненты:
Наличие в области достаточного количества “базовых” микроорганизмов, несущих основную часть генома будущего паразитического микроба;
Наличие недостающей части генома, способной перейти в геном “базового” микроорганизма;
Наличие высокочувствительных и высокочувствительных животных, служащих “горючим материалом” для“горящей искры”.
При совпадении всех вышеперечисленных условий запускается первая стадия цикла развития популяции микробов.
Накопление микробов в организме конкретного животного и на территории царства в целом. Предполагая, что плазмиды могут представлять собой трансформированные геномы фагов или других вирусов, в сочетании с геномом микроба - реципиента, нужно было бы только найти доноров плазмид чумного микроба. Если процесс эволюции чумного микроба завершился давным - давно, а вирусов, предшественников плазмид Y. pestis, в настоящее время не существует, мы не сможем найти их в царствах возбудителя чумы. Но если принять гипотезу А. И. Дятлова о Y. в современный период, фактически с каждой вспышкой, целенаправленный поиск приведет к обнаружению всех компонентов генетического материала и “основного” микроба, которые в сочетании образуют возбудителя чумы. Теоретически не важно, где они встречаются и сливаются – во внешней среде, откуда комбинированный продукт попадает в организм восприимчивого животного (теллурическая чума) /2,3/ или основной микроб уже живет в организме восприимчивого животного, но он не вреден без дополнительного компонента. Это может быть, например, микроб, входящий в нормальную микрофлору кишечного тракта, полости рта или носоглотки ("фарингеальная чума” Бальтазара). Когда недостающий компонент вторгается в организм извне, геномы объединяются, что приводит к процессу накопления вирулентного микроба. Реальность “бинарного” механизма образования, в частности, чумного микроба, может быть подтверждена его экспериментальным воспроизведением, например, на компонентах микрофлоры кишечного содержимого модельных животных, обязательно, высокочувствительных к данной инфекции. Интересные результаты могут быть получены также в экспериментах по трансформации микробов - компонентов нормальной микрофлоры кишечника - фрагментами нуклеиновых кислот, определяющими вирулентность, с использованием свободных от микробов, определенно высокочувствительных животных. Естественно, что все перечисленные выше процессы весьма вероятны, а для их реализации необходимо совпадение конкретных условий. Основным из них, является количество и концентрация на территории предстоящего участка компонента “основного” генома микроба, количество и концентрация второй части генетического материала, определяющего вирулентность, а также количество и концентрация “легковоспламеняющегося материала”, т. е. высокочувствительных и высокочувствительных животных (группа I по туляремии).
Здравый смысл подсказывает , что если из трех компонентов два компонента совпадают, т. е. “основной” микроб находится внутри организма чувствительного животного, вероятность генерации вирулентного микроба возрастает.
Из приведенных выше рассуждений можно сделать теоретические и практические выводы. Теоретические: Популяцию микроорганизмов в определенной области, можно рассматривать, как единый “многоклеточный организм” с качественно иным, по сравнению с отдельной клеткой, уровнем взаимодействия с окружающей средой. Для патогенных микроорганизмов средой может быть хозяин - макроорганизм. Для обсуждения предлагается концепция двухэтапной стратегии развития микробной популяции.
Первый этап - это увеличение численности микроорганизмов на территории конкретного царства или в организме животного / человека - хозяина.
Второй этап - это переход, также с использованием низкочувствительного хоста, в новую область. При благоприятных условиях, на новой территории начинается рост численности населения, обеспечивающий очередной скачок. При невозможности реализации первого и второго этапов, нельзя исключать диссоциацию патогенного микроорганизма на непатогенный (единичные) или разделение его генома на отдельные фрагменты, оставшиеся в клетках сапрофитов во внешней среде или в клетках нормальной микрофлоры высокочувствительного хозяина - макроорганизма.
В качестве практической реализации представленной концепции стратегии микробной популяции можно рассматривать тот факт, что она должна учитываться при разработке средств и методов профилактики, диагностики и лечения инфекционных заболеваний.
Рис. 1. Соответствие кривой роста микробных культур на искусственной питательной среде циклам в естественной экосистеме и в организме человека.
В питательной среде: Инокуляция среды Лаг-фазы (латентной)
Логарифмическая фаза
Полномасштабная стационарная фаза
Логарифмическая фаза смерти
Сохранение Фазы инфекционного заболевания
Инфекция восприимчивого организма
Период инкубации
Продромальное развитие клинического проявления
Полномасштабное клиническое проявление Криз (лизис)
Реконвалесценция
Превращение в хронический процесс, состояние носителя Фазы в переходной области:
Сохранение, некультивированные формы
Инициация эпидемического (эпизоотического) процесса (микроплощадки) Вспышка, логарифмическая фаза накопления
Переход популяции микробов к популяции менее чувствительных животных Снижение численности микробов на территории (микроплощадках)
Сохранение, передача на новую территорию
Рис. 2. Полный цикл существования в природе вида (популяции) патогенных микроорганизмов, например Yersinia pestis, Francisella tularensis, Burkholderia mallei. Вспышка в межэпизоотический период (микроплощадки инфекции) Эпизоотические эпидемиипрофитная (почвенная) комменсальная (глоточная) вирулентная форма “основного генома”, например в глотке высоковосприимчивого животного, в почве, в высоких пороговых концентрациях
Источник дополнительной части генома (плазмида, фаг, неизвестный микроорганизм), некультивированная формаккумуляция численности (части популяционного организма) Туш восприимчивых животных, их фекальная аккумуляция на территории генетического материала и его частичная консервация в некультивированной форме
Механизм передачи консервации менее восприимчивых животных, человека, перевозка высокочувствительных животных (1-2%)
Передача материала в новый реальный новый инфекционный очаг в новом реальном носителе
Заражение высокочувствительных животных, накопление (рост) популяции Вовлечение пути передачи.
REFERENCES
A.P. Gintsburg, Yu.M. Romanova// Microbiology Journal, 1997, N.3, pp. 116-121
I.V. Domaradsky “Plague: Modern state, hypotheses, problems”, Saratov, 1993 I.V. Domaradsky// Microbiology Journal, N. 4, pp. 16-20, 1997.
A.I. Diatlov. Evolutionary aspects in the plague natural nidality. – Stavropol, 1989.
A.I. Diatlov// Microbiology Journal, N. 6, pp. 113-115, 1999.
S.A. Lebedeva, A.V. Rakin, V.C. Ivanova et al.// Natural-nidal infections in Russia: Abstracts of the All-Russia scientific and practical conference “Modern epidemiology, diagnostics, tactics of the population protection”, pp. 11-12, Omsk, 1998.
V.Yu. Litvin// Microbiology Journal, N. 4, pp. 26-31, 1997.
V.Yu. Litvin// Natural-nidalinfections in Russia: Abstracts of the All-Russia scientific and practical conference “Modern epidemiology, diagnostics, tactics of the population protection”, pp. 20-21, Omsk, 1998.
V.Yu. Litvin, A.P. Gintsburg// Microbiology Journal, N. 4, pp. 63-72, 2002.
V.V. Prozorovsky, I.V. Pakovskaya, L.G. Gorina et al.// Microbiology Journal, N. 4, pp. 47-51, 1997.
V.I. Pushkareva, E.N. Emelyanenko, V.Yu. Litvin et al.// Microbiology Journal, N. 3, pp. 3-6, 1997.
L.V. Romanova, B.N. Mishankin, N.L. Pichurina et al.// Microbiology Journal, N. 2, pp. 11-15, 2000
O.M. Romanova, N.V. Alekseeva, A.L., A.L. Gintsburg// Microbiology Journal, N. 2, p. 26, 2000
V.M. Ruchko, L.I. Makhlai, V.A. Maksimov et al.// Problems of Especially Dangerous Diseases, v. 80, pp. 157-163, Saratov, 2000.
Yu.G. Suchkov, I.V. Khudiakov, E.N. Emelyanenko et al.// Microbiology Journal, N. 4, pp. 42-46, 1997
A.A. Filippov// Problems of Especially Dangerous Diseases, v. 80, pp. 71-88, Saratov, 2000.
B.K. Cherkassky. Manual on General Epidemiology,2001.
B.L. Cherkassky. Epidemiology and Prevention of Anthrax. M., 2002.
T. Johanson, K. Hagg, I. Golovlev et al. The 3rdInternational Conference on Tularemia. P. 22, Umea, 2000.
I.V. Romanova, B.N. Mishankin, N.L. Pichurina. The 3rdInternational Conference on Tularemia. P. 25, Umea, 2000
Новости, профилактика и раннее лечение ОРВИ:
Телеграм канал COVID-19 NOTE https://t.me/covidnotes