2
2. Транспорт веществ через биологические мембраны. Пассивный транспорт. Ионные каналы. Аквапорины. Активный транспорт веществ через мембраны. Первично-активный транспорт. Строение транспортных АТФ-аз. Вторичноактивный транспорт. Эндоцитоз и экзоцитоз: биохимические механизмы.
Транспорт веществ через биологические мембраны:
Весь транспорт делится на активный и пассивный:
Пассивный:
- Осмос
- Простая диффузия
- Облегченная диффузия
Активный:
- Первично-активный транспорт
- Вторично-активный транспорт
- Везикулярный транспорт: эндоцитоз (пино-, фагоцитоз) и экзоцитоз
Пассивный транспорт:
1) Осмос - движение жидкости, газов из области с меньшей концентрацией в большую
2) Простая диффузия - просто перемещение частиц туда-сюда в следствии их движения (без транспортных белков, просто через билипидный слой)
3) Облегченная диффузия - транспорт молекул через мембрану при помощи специальных транспортных белков
Ионные каналы
Это порообразующие белки, осуществляют облегченную диффузию. Ну тут можно просто шизу воткнуть, что есть независимые, потенциал-зависимые, лиганд-зависимые, механочувствительные, также про работу створок канала
Аквапорины
- Ионные каналы, отвечающие за транспорт воды.
- Избирательно пропускают молекулы воды, позволяя ей поступать в клетку и покидать её, в то же время препятствуя протоку ионов и других растворимых веществ.
- Регулируются фосфорилированием, которое приводит к большей активности аквапорина
Активный транспорт веществ через мембраны. Первично-активный транспорт:
Здесь уже участвуют ионные насосы, а не каналы.
Насосы переносят вещества против градиента концентрации за счет энергии гидролиза АТФ
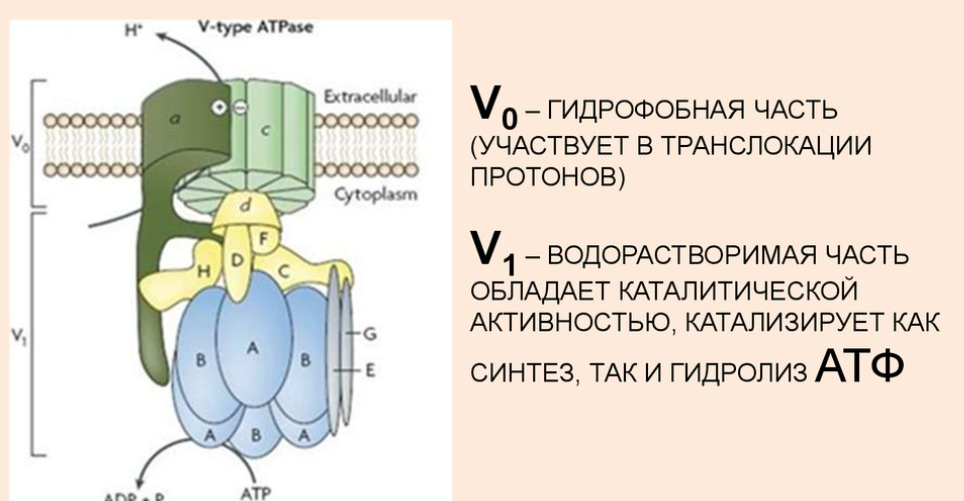
Строение транспортных АТФ-аз:
АТФазы - ферменты, гидролазы. Отсоединяют от АТФ 1 или 2 остатка фосфорной кислоты с высвобождением энергии.
Переносят протоны

Вторичноактивный транспорт.
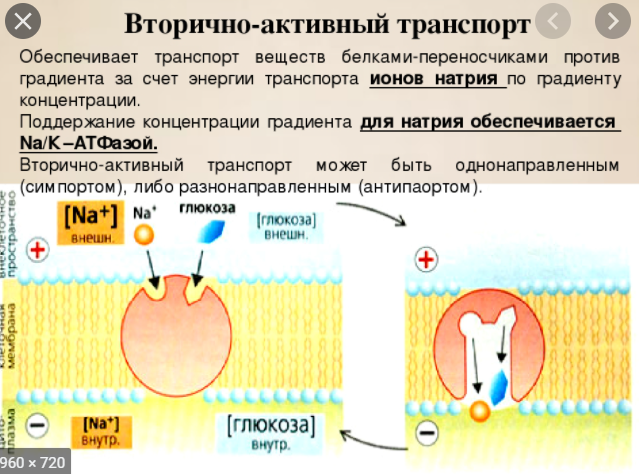
Эндоцитоз и экзоцитоз: биохимические механизмы
Эндоцитоз - перенос веществ из среды в клетку вместе с частью плазматической мембраны, путем образования мембраной пузырьков.
Обычно клетки формируют пузырьки постоянно, ритмично и постоянно что-то захватывают из внеклеточной жидкости, но иногда эндоцитоз индуцируется веществами, контактирующими с клеткой. Например, рецепто ры плазматической мембраны гепатоцитов улавливают многие гликопротеины плазмы крови, а затем эти белки эндоцитируются.
Эндоцитоз:
- процесс, нуждающийся в энергии
- источником последней служит АТФ
- потребителем энергии являются сокращающиеся микрофиламенты
- присоединение лиганда к рецепторам изменяет их конформацию => после чего к ним присоединяются микрофибриллы => их сокращение приводит к впячиванию мембраны и отделению пузырька
Экзоцитоз:
Есть два основных типа экзоцитоза в эукариот: регулируемые и нерегулируемые (или составные). Регулируемый экзоцитоз, который требует внешнего сигнала, используется для высвобождения нейротрансмиттеров и секреции гормонов. В отличие от регулируемого экзоцитоза, составной экзоцитоз осуществляется всеми клетками. Клетки используют составной экзоцитоз для высвобождения компонентов внеклеточной матрицы или включения белков в плазменную мембрану.
Моторные белки активно перемещают пузырьки по цитоскелетам микротрубочек и нитей. Вторым шагом является привязка пузырьков, в которой пузырьки связываются с плазменной мембраной. На третьем этапе, стыковке пузырьков, мембрана пузырьков прикрепляется к плазменной мембране, и две мембраны начинают сливаться.
Четвертый шаг, праймирование пузырьков, происходит только при регулируемом экзоцитозе. Праймирование включает в себя изменения, происходящие после привязки пузырька, но прежде чем он выпускает свое содержимое. Праймирование готовит пузырьки для синтеза с плазменной мембраной.
Пятый шаг - слияние пузырьков. Оно может быть полным или по схеме "поцеловал-и-убежал". При полном слиянии пузырьки полностью разрушаются и становятся частью плазменной мембраны, изгоняя их содержимое из клетки в процессе.
4. Катаболизм и анаболизм. Общая схема обмена веществ и энергии в организме человека. Специфические и общие пути катаболизма.
Анаболизм
Анаболизм – это биосинтез белков, полисахаридов, липидов, нуклеиновых кислот и других макромолекул из малых молекул-предшественников. Поскольку он сопровождается усложнением структуры, то требует затрат энергии. Источником такой энергии является энергия АТФ.
Катаболизм
Катаболизм – расщепление и окисление сложных органических молекул до более простых конечных продуктов. Оно сопровождается высвобождением энергии, заключенной в сложной структуре веществ. Большая часть высвобожденной энергии рассеивается в виде тепла. Меньшая часть этой энергии "перехватывается" коферментами окислительных реакций НАД и ФАД, некоторая часть сразу используется для синтеза АТФ.
Общая схема обмена веществ и энергии в организме человека:
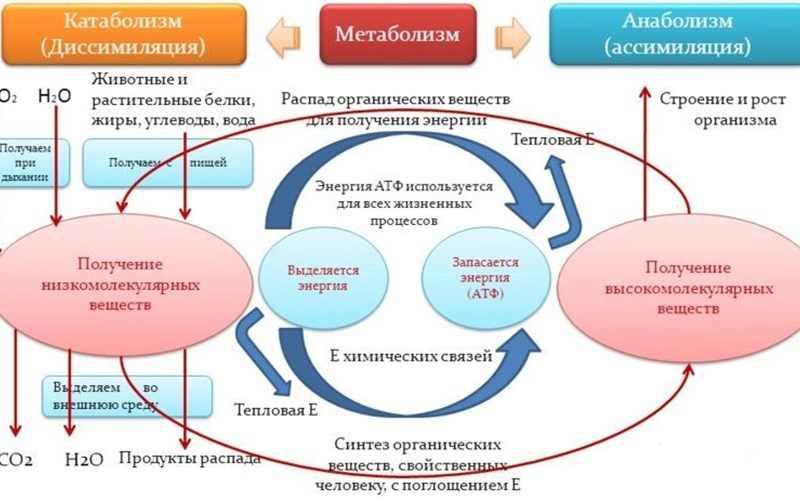
Специфические и общие пути катаболизма:
В катаболизме различают три стадии:
1) Полимеры превращаются в мономеры (белки – в аминокислоты, углеводы в моносахариды, липиды – в глицерол и жирные кислоты). Химическая энергия при этом рассеивается в виде тепла.
2) Мономеры превращаются в общие продукты, в подавляющем большинстве в ацетил-КоА. Химическая энергия частично рассеивается в виде тепла, частично накапливается в виде восстановленных коферментных форм (НАДН, ФАДН2), частично запасается в макроэргических связях АТФ (субстратное фосфорилирование).
1-ая и 2-ая стадии катаболизма относятся к специфическим путям, которые уникальны для метаболизма белков, липидов и углеводов.
3) Заключительный этап катаболизма, сводится к окислению ацетил-КоА до СО2 и Н2О в реакциях цикла трикарбоновых кислот (цикла Кребса) – общий путь катаболизма. Окислительные реакции общего пути катаболизма сопряжены с цепью тканевого дыхания. При этом энергия (40-45%) запасается в виде АТФ (окислительное фосфорилирование).
6. Цикл трикарбоновых кислот. Общая схема, энергетическое значение
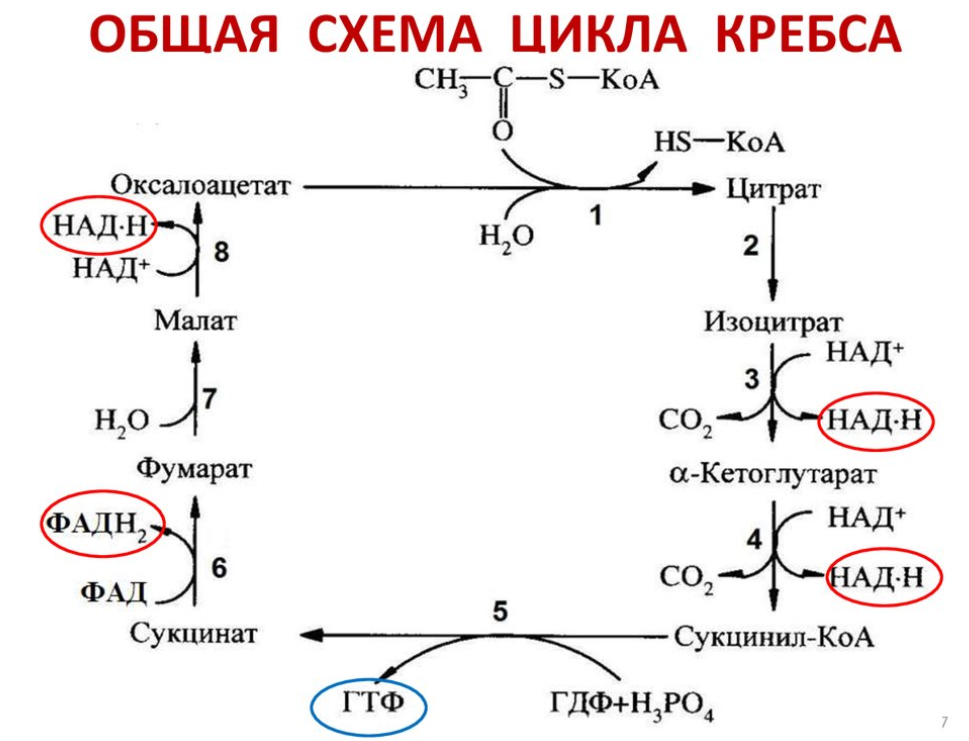
Энергетическое значение:
1) Образуется ГТФ = 1АТФ
2) 3НАДН и 1ФАДН2 - источники водорода для дыхательной цепи, т.е. окислительного фосфорилирования
8. Регуляция цикла трикарбоновых кислот. Анаболические функции цикла трикарбоновых кислот. Анаплеротические реакции
Регуляция цикла трикарбоновых кислот
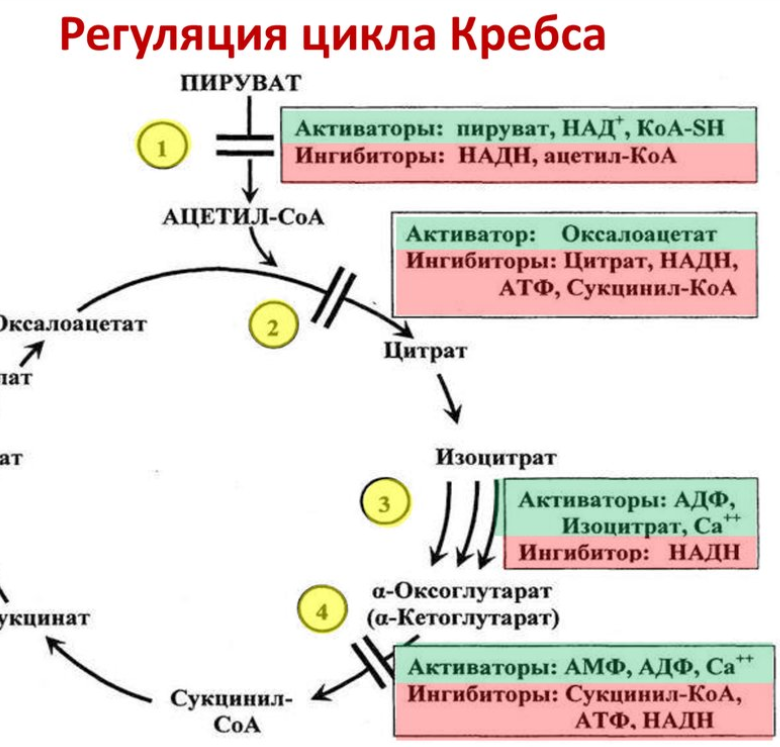
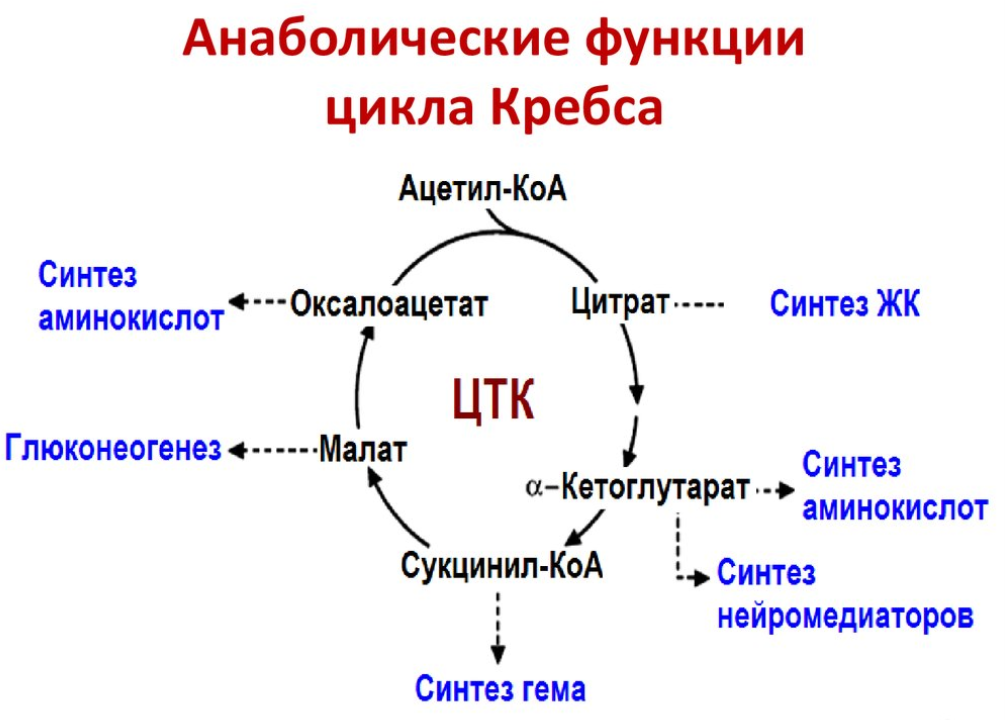
Анаплеротические реакции
Анаплеротические (пополняющие) реакции – специальные ферментативные реакции, обеспечивающие пополнение пула промежуточных продуктов цикла лимонной кислоты.
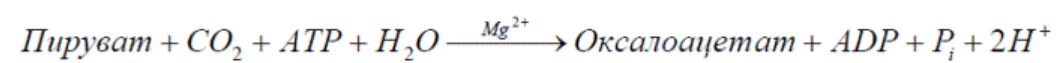
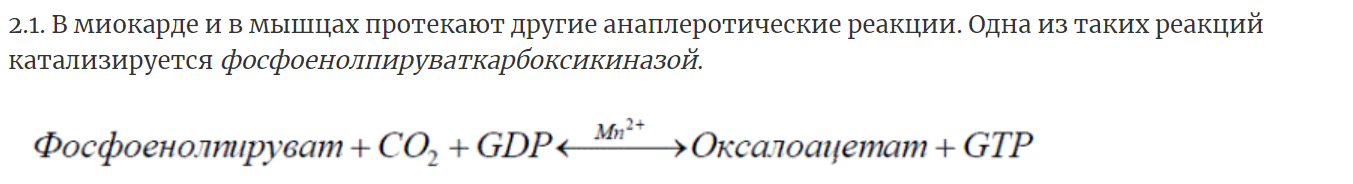

10. Пути синтеза АТФ: субстратное и окислительное фосфорилирование. Примеры реакций субстратного фосфорилирования.
Окислительное фосфорилирование – многоэтапный процесс во внутренней мембране митохондрий, происходит окисление восстановленных НАДН и ФАДН2 и сопровождающийся синтезом АТФ.
Субстратное фосфорилирование – фосфорилирование АДФ до АТФ. Передача макроэргического фосфата или энергии макроэргической связи на АДФ.
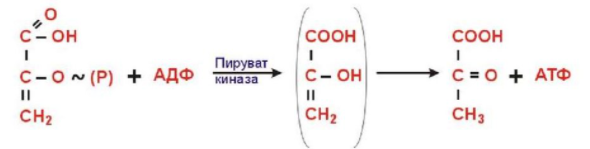
Фосфоенолпируват + АДФ -> енольная форма пирувата -> пируват
12. Состав, структура и номенклатура дыхательных комплексов и других компонентов цепи переноса электронов, их локализация и функции во внутренней мембране митохондрий. Строение АТФ-синтазы. Механизм функционирования.
Всего цепь переноса электронов включает в себя разнообразные белки, которые организованы в 4 больших мембраносвязанных мульферментных комплекса. Также существует еще один комплекс, участвующий не в переносе электронов, а синтезирующий АТФ.
Состав, структура и номенклатура дыхательных комплексов
1 комплекс. НАДН-КоQ-оксидоредуктаза (НАДН-дегидрогеназа)
Структура:
ФМН, 42 белковых молекулы, из них не менее 6 железосерных белков
Функция
- Принимает электроны от НАДН и передает их на коэнзим Q (убихинон).
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
2 комплекс. ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует, его выделение условно. К нему относятся ФАД-зависимые ферменты, расположенные на внутренней мембране:
- ацил-SКоА-дегидрогеназа (β-окисление жирных кислот)
- сукцинатдегидрогеназа (цикл трикарбоновых кислот)
- фосфат-дегидрогеназа (челночный механизм переноса атомов водорода)
Функция
- Восстановление ФАД в окислительно-восстановительных реакциях.
- Обеспечение передачи электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q
3 комплекс. КоQ-цитохром c-оксидоредуктаза (цитохром с редуктаза)
Строение: молекулы цитохрома b и цитохрома c1, железо-серные белки. 2 мономера с 11 полипептидными цепями.
Функция
- Принимает электроны от коэнзима Q и передает их на цитохром с.
- Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
4 комплекс. Цитохром с-кислород-оксидоредуктаза (цитохромоксидаза)
Строение: 13 субъединиц, включает цитохромы а и а3
Функция
- Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
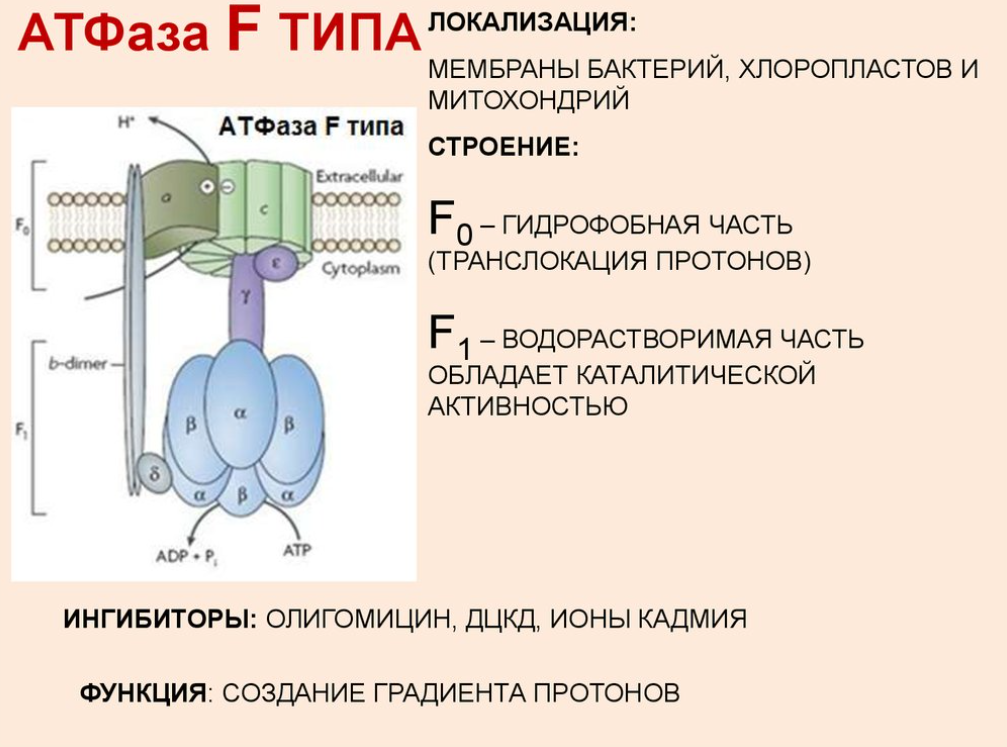
5 комплекс. АТФ-синтаза
Строение: множество белковых цепей, разделенных на 2 группы:
- cубъединица F0, каналообразующая, по ней выкачанные наружу протоны водорода устремляются в матрикс
- cубъединица F1, каталитическая, используя энергию протонов, синтезирует АТФ
Механизм работы АТФ-синтазы получил название вращательный катализ:
- синтез АТФ происходит за счет энергии, высвобождающейся при прохождении протонов через каналы комплекса Fo
- протоны из межмембранного пространства достигают своих центров связывания на с-субъединицах через входной полуканал и прикрепляются к аспартату, нейтрализуя его отрицательный заряд -> лишенная заряда с-субъединица меняет свою конформацию и принуждает Fo-комплекс вращаться вокруг своей оси, доставляя протоны к другому полуканалу, который направлен уже в матрикс митохондрий -> поскольку концентрация ионов H+ в матриксе низка, то они легко отрываются от аспартата и уходят внутрь, а оставшийся, уже заряженный, аспартат не позволяет комплексу вращаться в обратном направлении.
-при связывании ионов H+ происходит поворот не только комплекса c-субъединиц Fo-компонента, но и жестко связанной с ним γ-субъединицы F1-компонента -> стержень γ–субъединицы проворачивается и при каждом повороте на 120° она поочередно вступает в контакт с каталитическими β-субъединицами, что меняет их функционирование -> при каждом обороте γ-субъединицы на 360º синтезируются три молекулы АТФ.
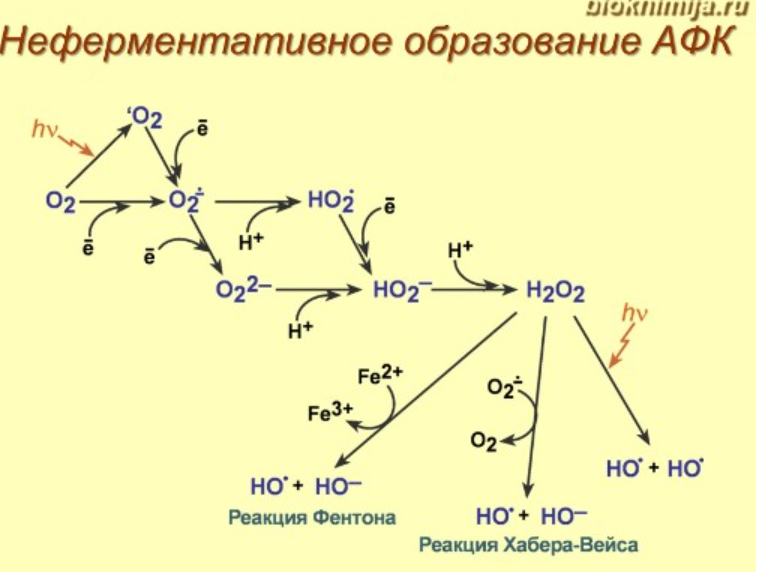
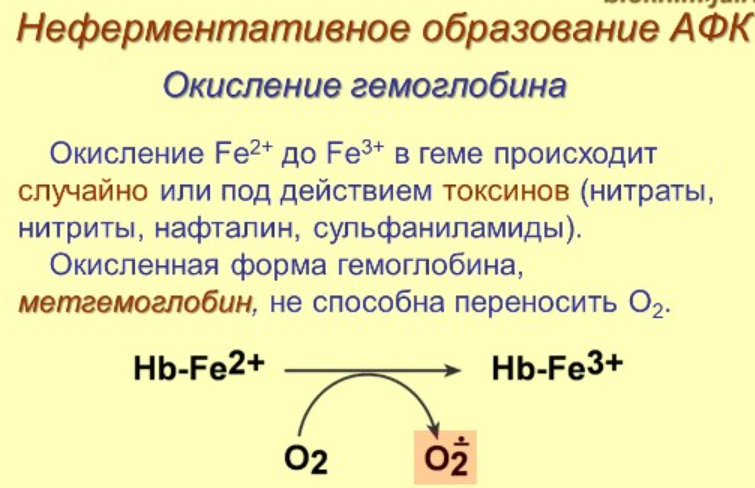
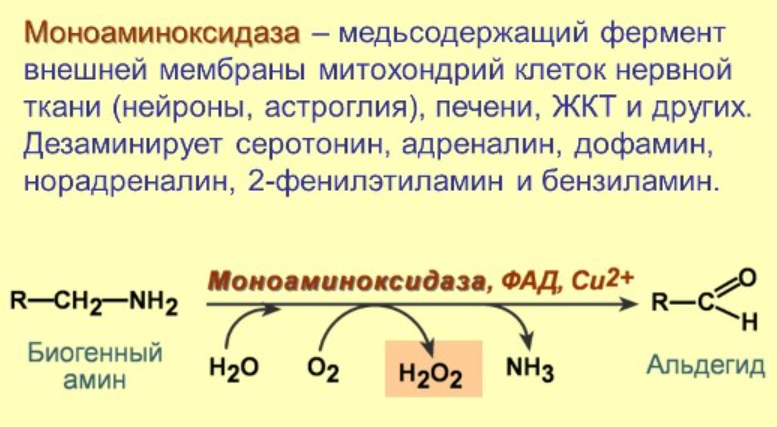
14. Активные формы кислорода и азота. Неферментативные и ферментативные пути образования. Перекисное окисление липидов: механизм процесса, продукты. Первичные и вторичные механизмы повреждающего действия окислительного стресса. Перекисная теория гибели клетки
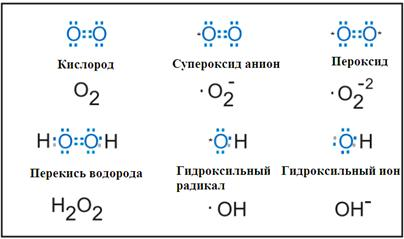
Активные формы азота
Пероксинитрит анион - ONOO-
См. тетрадь
Ферментативные пути образования:
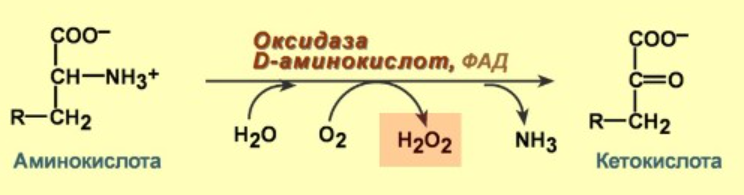
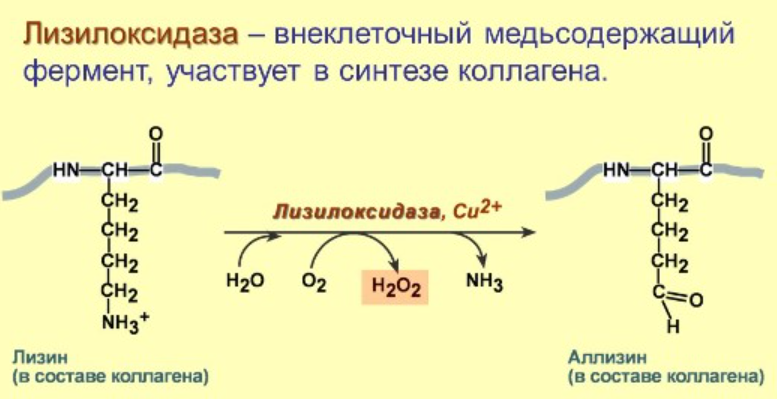
Перекисное окисление липидов: механизм процесса, продукты
При окислительном стрессе лидирует окисление жирнокислотных остатков в фосфолипидах мембран. Это снижает их гидрофобность и нарушает устойчивость мембран, изменяет работу мембраносвязанных ферментов, повышает проницаемость мембран для ионов.
Перекисное окисление липидов включает в себя несколько стадий:
- Инициация - атакуется метиленовая группа, расположенная между двойными связями, и выбивается атом водорода, восстанавливающий гидроксил-радикал до воды. Далее в жирной кислоте происходит перестановка двойной связи, смещение радикальной группы и взаимодействие ее с кислородом. В результате образуется липопероксильный радикал
- Развитие - дальнейшее взаимодействие полученного липопероксильного радикала с соседними жирными кислотами приводит к его нейтрализации и появлению новых липоперекисных радикалов, т.е. к развитию линейной цепной реакции с появлением новых окисленных жирных кислот.
- Разветвление - кроме линейного развития, может происходить ветвление реакции за счет получения гидроперекисью электронов от каких-либо металлов или при воздействии излучения
- Обрыв цепи - обрыв цепной реакции происходит при взаимодействии радикалов друг с другом или в реакции с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в довольно стабильную окисленную форм
Продукты:
Первичные - гидроперекиси жирных кислот
Вторичные (от распада гидроперекиси) - спирты, кетоны, альдегиды. Запомнить - малоновый диальдегид (наиболее реакционноспособный)
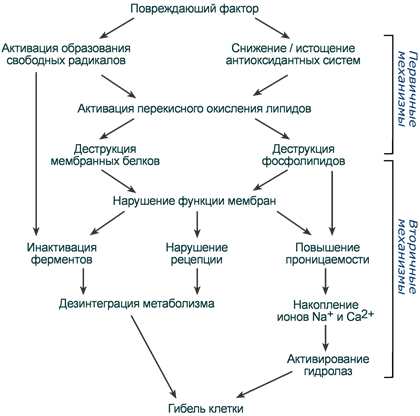
Первичные и вторичные механизмы повреждающего действия окислительного стресса
Окислительный стресс - избыточная продукция эндогенных АФК или из-за большой концентрации экзогенных АФК
Первичные м-мы:
Процессы окисления непосредственно в местах своего образования – митохондриях, клеточной мембране, мембранах ЭПР. Под влиянием свободных радикалов происходит окисление молекул – ядерной и митохондриальной ДНК, РНК, белков, фосфолипидов и гликозаминогликанов
Вторичные м-мы:
Деструкция мембран, внутриклеточных структур, органелл
Свободнорадикальная теория старения утверждает, что старение происходит из-за накопления повреждений в клетках, нанесённых свободными радикалами с течением времени
Свободнорадикальная митохондриальная теория старения, представляющая собой модификацию свободнорадикальной теории старения, утверждает, что старение у людей и животных обусловлено накоплением повреждений в митохондриях и митохондриальной ДНК[
16. Физиологическое значение свободно-радикального окисления. Свободнорадикальное окисление, фагоцитоз и воспаление. Свободнорадикальный механизм антимикробной защиты грудного молока. Свободнорадикальное окисление при патологии сердечно-сосудистой системы.
Физиологическая роль свободных радикалов достаточно велика. Большая часть свободных радикалов генерируются фагоцитами, Т-лимфоцитами при воспалительных реакциях и выполняют защитную роль, лизируя патогенные микроорганизмы, мутировавшие (раковые) клетки.
Важнейшую физиологическую роль выполняет генерируемый в эндотелии сосудов оксид азота NO (эндотелий-зависимый релаксирующий фактор), обеспечивающий расслабление гладкой мускулатуры сосудистой стенки и регулирующий уровень артериального давления, коронарный и органный кровоток, а также предотвращающий агрегацию тромбоцитов
Свободнорадикальное окисление, фагоцитоз и воспаление
Фагоцитоз – это активное поглощение и разрушение клетками твердых частиц, чуждых для организма (микроорганизмы, вирусы, иммунные комплексы антиген-антитело, простейшие, гельминты, отмершие клетки, твердые неорганические частицы)
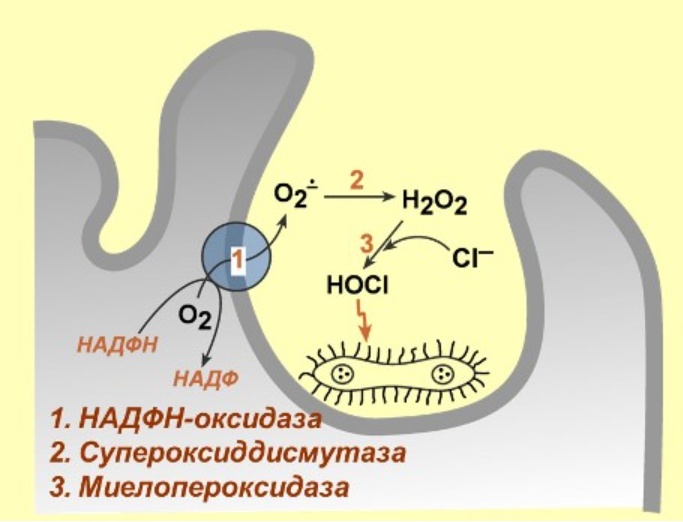
Свободнорадикальный механизм антимикробной защиты грудного молока


Свободнорадикальное окисление при патологии сердечно-сосудистой системы
При гипоксии (ишемии) миокарда окислительные процессы в митохондриях кардиомиоцитов нарушаются (как бы не доходят до конца), в результате чего накапливаются промежуточные метаболиты цикла Кребса, крайне легко подверженные восстановлению с образованием свободных радикалов и перекисных соединений, угнетающих систему антиоксидантной защиты. В конечном итоге создается парадоксальная ситуация — уменьшение кислорода в клетке приводит к увеличению кислородных радикалов. Развивающаяся после каждого эпизода транзиторной ишемии реперфузия миокарда также сопровождается значительной активацией (в сотни раз) свободнорадикальных процессов и выбросом липопероксидов в кровоток.
Велика роль свободнорадикального окисления (СРО) в патогенезе нестабильной стенокардии и инфаркта миокарда. Локальная активация СРО в зоне ишемии и накопление продуктов деградации свободных радикалов стимулируют свертываемость крови, увеличивают ее вязкость, усиливают агрегацию и адгезию форменных элементов крови. Высокая концентрация пероксидов ускоряет дегенерацию NO с образованием пероксинитрита — крайне цитотоксичного соединения: О2– +NOо ->ОNOO–. Ускоренный распад эндотелиального NO стимулирует ангиоспазм, а окисление экзогенного NO, образующегося в результате метаболизма потребляемых больным нитропрепаратов, уменьшает их терапевтическую эффективность. Кроме того, свободные радикалы модифицируют эндотелиальные NO-рецепторы, уменьшая их чувствительность, а также оказывают непосредственное повреждающее действие на кардиомиоциты. Указанные процессы усугубляют ишемию, оказывают аритмогенный эффект, способствуют распространению зоны некроза и повреждения.
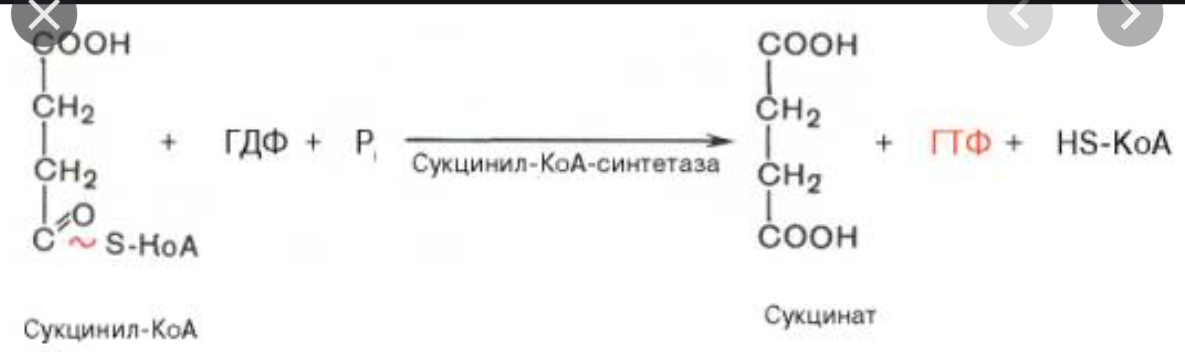
7. Последовательность реакций цикла трикарбоновых кислот. Реакции дегидрирования. Субстратное фосфорилирование.
Дегидрирование
Дегидрирование и декарбоксилирование изоцитрата НАД+ зависимой изоцитратдегидрогеназой с образованием a-кетоглутарата:
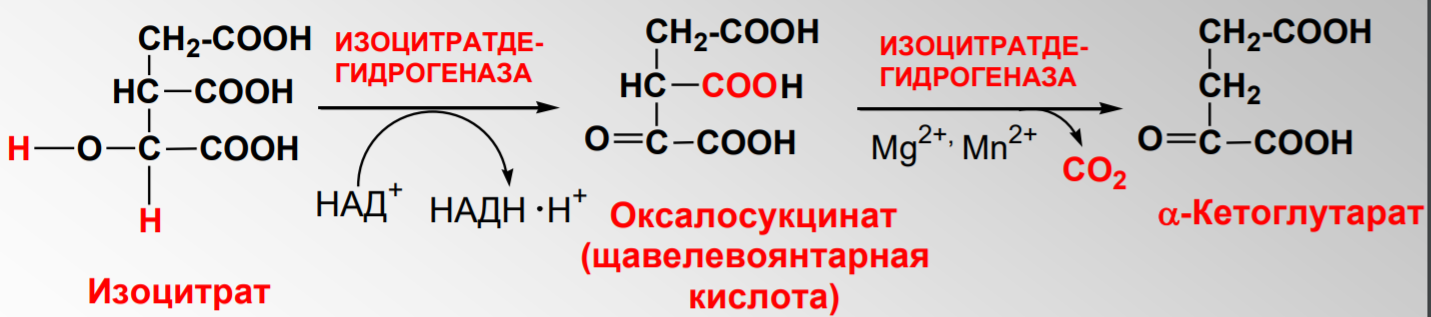
Дегидрирование сукцината до фумарата:
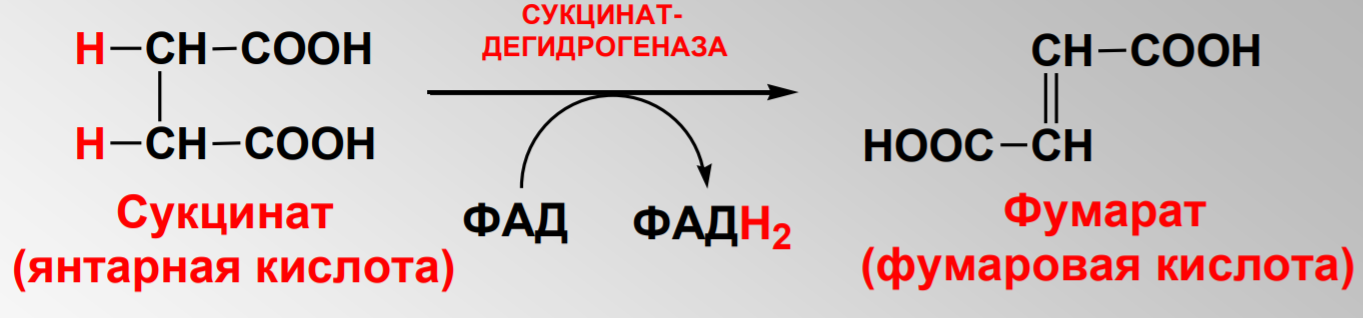
Дегидрирование малата
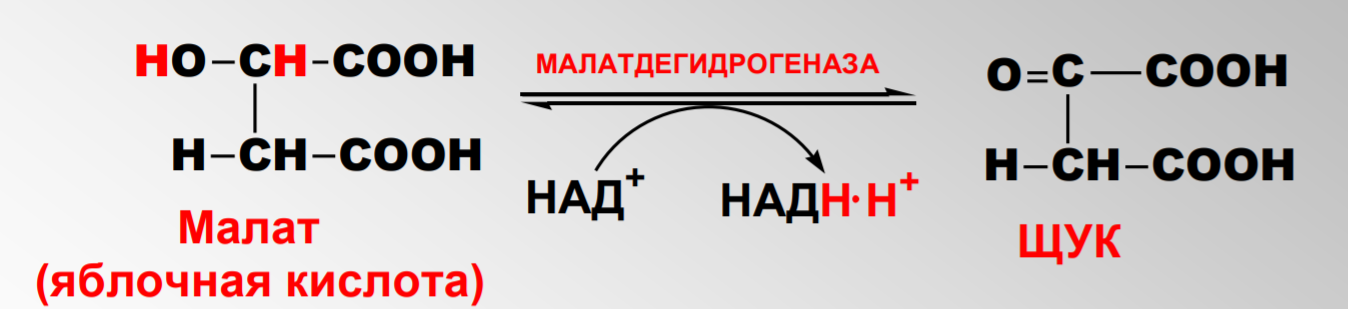
Окислительное фосфорилирование