Хуй
2. Сложные белки. Классификация, строение, примеры. Строение и функции миоглобина
Строение:
У сложных белков, как раз таки кроме белковой цепи (из амк) имеется небелковая часть - лиганд, связанный с белковой цепью. Если лиганд имеет структурную или функциональную нагрузку (или вообще и то и другое сразу) то он переименовывается в простетическую группу.
Классификация:
- нуклеопротеины - белки с НК. Например: рибосомы, хроматин, вирусы. В хроматине - НК связана с белками: гистоновыми (отвечают за начальную упаковку) и негистоновыми.
- фосфопротеины - белки с фосфатной группой. Она связывается с пептидной цепью через остатки тирозина, серина и треонина (тех аминокислот, которые содержат ОН-группу). Например, ферменты: гликогенсинтаза и гликогенфосфорилаза.
На всякий: в клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную (неактивную и активную)
- металлопротеины - с катионами металлов, катион связан с функциональными группами (-COOH или -NH2) амк белка. Примеры: ферритин(запас железа), альфа-амилаза.
- липопротеины - белок с простетической группой в виде липида. В составе могут быть: свободные жирные кислоты, нейтральные жиры, фосфолипиды, холестериды. Хуй знает нужно будет гандонам или нет, но:
Различают четыре основных класса лиипопротеинов:
-липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП),
-липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП),
-липопротеины очень низкой плотности (ЛПОНП)
-хиломикроны (ХМ)
- хромопротеины - с окрашенными простетическими группами. Например: гемопротеины (простетическая группа - гем), ретинальпротеины (простетическая группа - витамин А), флавопротеины (простетическая группа - витамин В2), кобамидпротеины (простетическая группа - витамин В12)
- гликопротеины - содержат углеводный компонент. А вообще, выделяют 2 подкласса белков с углеводами: гликопротеины и протеогликаны. Основное отличие в том, что у гликопротеинов: 15-20% углеводов в составе, у протеогликанов: 80-85% (по названию походу в конце стоит то, чего больше)
Миоглобин: Строение:
Молекула миоглобина образована глобулярным белком с единичной полипептидной цепью и железосодержащим гемом, молекула сходна по строению и функциям с гемоглобином крови.
Функции:
Депо кислорода в мышцах, использование только при сильном уменьшении рО2. Т.о., миоглобин остается оксигенированным до того момента, пока количество кислорода в клетке не снизится до предельно малых величин.
3. Строение и функции гемоглобина. T- и R-формы гемоглобина. Аллостерические эффекты гемоглобина: кооперативный эффект, эффект Бора, эффект 2,3-дифосфоглицерата. Механизмы, физиологическое значение. Особенности фетального гемоглобина. Понятие о гемоглобинопатиях. Серповидно-клеточная анемия. Талассемия.
Строение и функции гемоглобина.
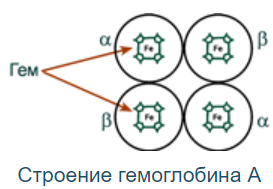
Строение:
Гемоглобин - белок с 4 субъединицами, содержащими гем (картинка). Между собой протомеры соединены связями:
- гидрофобными
- ионными
- водородными
Соединены контактными поверхностями, в десятках точек по принципу комплементарности (разноимённо заряженные группы, гидрофобные участки, неровности на поверхности белка).
T- и R-формы гемоглобина:
Дезоксиформа гемоглобина обозначается как Т-форма, напряженная, она обладает существенно более низким сродством к кислороду. Оксигенированная форма, или R-форма обладает высоким сродством к кислороду.
Кооперативный эффект:
Связывание О2 с одной из субъединиц T-формы приводит к локальным конформационным изменениям, которые ослабляют связь между субъединицами, облегчая присоединение последующих молекул кислорода и изменение других протомеров.
Эффект Бора:
Влияние рН на сродство гемоглобина к кислороду носит название эффекта Бора. При закислении среды сродство снижается, при защелачивании – повышается.
- При повышении концентрации протонов (закисление среды) в тканях возрастает освобождение кислорода из оксигемоглобина.
- В легких после удаления угольной кислоты (в виде СО2) из крови и одновременном увеличении концентрации кислорода высвобождаются ионы Н+ из гемоглобина
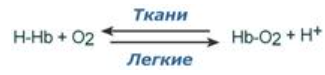
Изменение сродства гемоглобина к кислороду в тканях и в легких (при изменении концентрации ионов H+ и О2) обусловлено конформационными перестройками глобиновой части молекулы.
В ткани:
Молекула О2 отрывается от железа и ионы водорода присоединяются к остаткам гистидина, образуется H-Hb с низким сродством к кислороду.
В легких:
Поступающий в больших количествах кислород "вытесняет" ион водорода из связи с остатком гистидина гемоглобиновой молекулы.
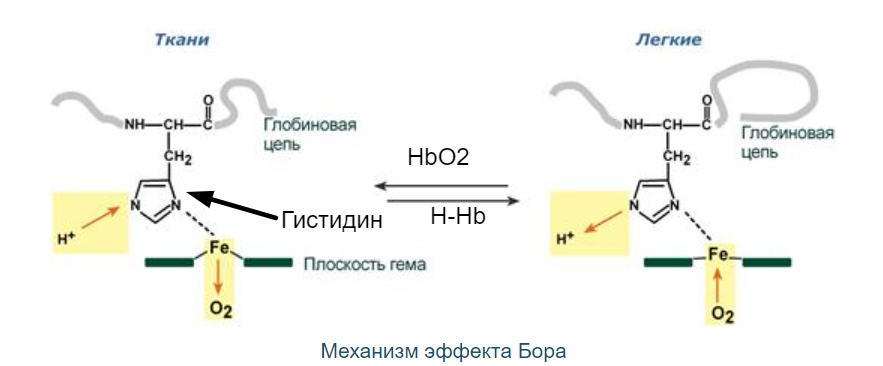
Роль 2,3-дифосфоглицерата
2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта.
2,3-Дифосфоглицерат располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями, образуя поперечный солевой мостик между атомами кислорода 2,3-дифосфоглицерата и аминогруппами обеих β-цепей
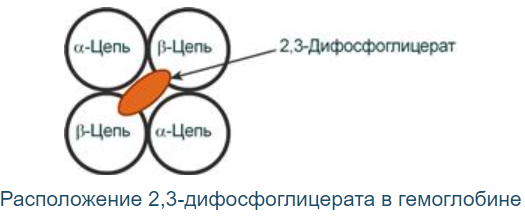
Функция:
Снижает сродство к гемоглобину (то есть отдача О2 в тканях возрастает в 2 раза). Имеет особенное значение при подъеме на высоту и при нехватке кислорода во вдыхаемом воздухе.
Особенности фетального гемоглобина:
- Состоит из 2х альфа и 2х гамма цепей (обычный взрослый из 2х альфа и 2х бета)
- Гемоглобин F обладает повышенным сродством к кислороду и позволяет сравнительно малому объёму крови плода выполнять кислородоснабжающие функции более эффективно
- гемоглобин F обладает меньшей стойкостью к разрушению и меньшей стабильностью в физиологически широком интервале pH и температур
Серповидно-клеточная анемия:
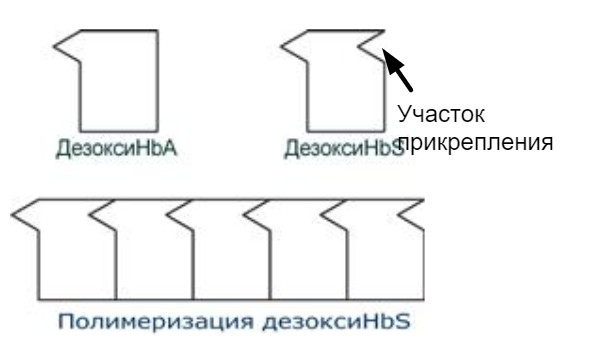
HbS – гемоглобин при серповидно-клеточной анемии. Возникает точковая мутация и в ДНК ЦТТ -> ЦАТ => в бета цепи появляется валин вместо глутамата. Из-за этого поверхность гемоглобина изменяется и появляется "участок прикрепления", за который могут зацепиться другие серповидно-клеточные гемоглобины, поэтому вся эта хуйня соединяется, полимеризуется, происходит деформация эритроцитов и при прохождении через капилляры - гемолиз.
Талассемия:
Для талассемий характерно снижение синтеза α-цепей гемоглобина (α-талассемия) или β-цепей (β-талассемия). Это приводит к нарушению эритропоэза, гемолизу и тяжелым анемиям.
4. Общая характеристика ферментов. Специфичность ферментов. Виды специфичности, примеры. Строение ферментов. Кофакторы и коферменты. Классификация и номенклатура ферментов.
Общая характеристика ферментов
В нормальных физиологических условиях биохимические реакции в организме протекают с высокими скоростями, что обеспечивается биологическими катализаторами белковой природы – ферментами.
Ферменты:
- Катализируют только энергетически возможные реакции
- Не изменяют направления реакции
- Не расходуются в процессе реакции
- Высокая скорость работы
- Высокая специфичность
- Мягкие условия работы (внутри клетки)
- Могут регулировать скорость реакции
- Скорость ферментативной реакции пропорциональна количеству фермента
Специфичность ферментов:
Специфичность - высокая избирательность действия ферментов, основана на комплементарности структуры субстрата и активного центра фермента.
Виды специфичности, примеры:
1) Стереоспецифичность - катализ только одного из стереоизомеров.
Пример:
- специфичность ферментов к L-амк
- специфичность к цис- и транс- изомерам (аспартаза реагирует только с транс-изомером – фумаровой кислотой, но не с малеиновой кислотой (цис-изомер))
2) Абсолютная специфичность - главное слово-абсолютная, т.е. один фермент производит катализ только одного вещества.
Пример:
- каталаза на Н2О2
- уреаза только мочевины
3) Групповая специфичность - катализ только при наличии определенной связи, хим.группы.
Пример:
- наличие пептидной связи для: пепсина, тромбина
- наличие α1,4-гликозидных связей в крахмале и гликогене для: α-амилазы слюнной и поджелудочной желез
- наличие ОН-группы для: алкогольдегидрогеназы
Строение ферментов:
Все ферменты являются белками и обладают всеми свойствами белков. Поэтому, подобно белкам, ферменты делятся на простые и сложные.
Простые:
Только из аминокислот: пепсин, трипсин, лизоцим
Сложные (холоферменты) состоят из 2х частей:
1) Апофермент - белковая часть из амк
2) Кофактор - небелковая часть:
а) Кофермент (НАД+, НАДФ+, ФАД)
б) Простетическая группа (гем, Fe2+, Ca2+, Zn2+)
Примеры:
Амилаза (кофактор - Са2+)
Лактатдегидрогеназа (Zn2+)
Сукцинатдегидрогеназа (содержит ФАД)
Классификация и номенклатура ферментов:
I класс – Оксидоредуктазы (ОВР)
название: донор электронов : акцептор электронов – оксидоредуктаза,
пример: алкоголь:НАД-оксидоредуктаза (алкогольдегидрогеназа)
II класс – Трансферазы (перенос различных групп от одного субстрата (донор) к другому (акцептор))
название: донор группы : акцептор группы – переносимая группа трансфераза
пример: АТФ:фруктозо-6-фосфат-фосфотрансфераза (Фосфофруктокиназа)
III класс – Гидролазы (разрыв внутримолекулярных связей в субстрате путем присоединения элементов Н2О)
название: гидролизуемый субстрат : отделяемая группа гидролаза
пример: триацилглицерол : ацилгидролаза (ТАГ-липаза)
IV класс – Лиазы (катализируют разрыв связей без присоединения Н2О)
название: расщепляемый субстрат : отделяемая группа – лиаза
пример: АТФ:дифосфат-лиаза (аденилатциклаза)
V класс – Изомеразы
название: субстрат – [ ] – реакция, где [ ] – обозначение, отражающее суть реакции
пример: α-D-глюкозо-1,6-фосфомутаза (фосфоглюкомутаза)
VI класс – Лигазы (синтез новых веществ)
название: субстрат 1 : субстрат 2 – лигаза
пример: L-глутамат:аммиак-лигаза (Глутаминсинтетаза)
7. Ингибирование ферментативной активности. Конкурентное и неконкурентное обратимое ингибирование. Необратимое ингибирование. Кинетические зависимости. Примеры.
Ингибирование ферментативной активности
Два основных пути ингибирования:
а) По прочности связывания: обратимое и необратимое
б) По отношению ингибитора к активному центру: конкурентное и неконкурентное
Конкурентное и неконкурентное обратимое ингибирование
При обратимом ингибировании происходит непрочное связывание ингибитора с функциональными группами фермента, вследствие чего активность постепенно восстанавливается.
Конкурентное ингибирование:
ингибитор похож на субстрат фермента => соперничество за акт.центр фермента => уменьшение связывания субстрата с ферментом и нарушению катализа
-при данном ингибировании максимальная скорость реакции остается вполне достижимой при создании высоких концентраций субстрата
Пример:
- ингибирование сукцинат-дегидрогеназы малонатом, который похож на сукцинат
- антиметаболиты (сульфаниламиды против бактерий)
Неконкурентное ингибирование:
Присоединением ингибитора не в активном центре, а в другом месте молекулы => изменение структуры акт.центра => связь акт.центра с субстратом невозможна
Пример:
- синильная кислота связывается с гемовым железом ферментов дыхательной цепи и блокирует клеточное дыхание
- ионы тяжелых металлов (Cu2+, Hg2+, Ag+) c SH- группами белков
!!!Изменение концентрации субстрата никак не влияет
Необратимое ингибирование:
Связывание или разрушение функциональных групп фермента, необходимых для его активности
Пример:
Диизопропилфторфосфат прочно и необратимо связывается с гидроксигруппой серина в активном центре фермента ацетилхолинэстеразы
8. Регуляторные белки. Ассоциация диссоциация. Примеры. Регуляция активности ферментов путем фосфорилирования-дефосфорилирования, частичным протеолизом.
Регуляторные белки:
- белки-рецепторы, воспринимающие сигнал
- белки-гормоны и другие вещества, передающие сигналы клеткам
Бля речь про белок-белковое взаимодействие: после влияния каких-либо факторов на специфичные белки изменяется активность этих белков, и они, в свою очередь, воздействуют на нужный фермент.
1)
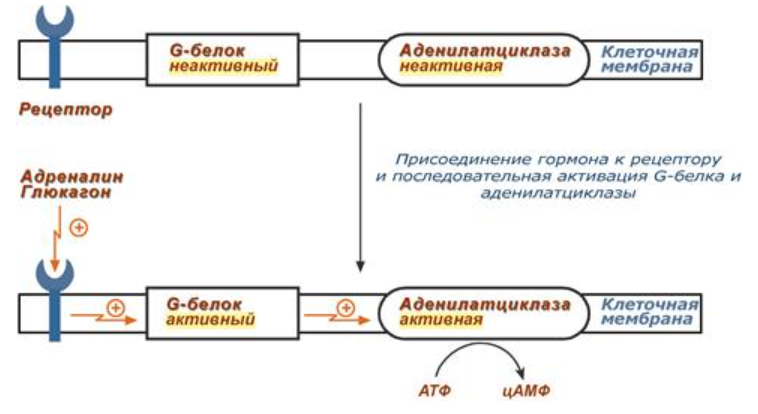
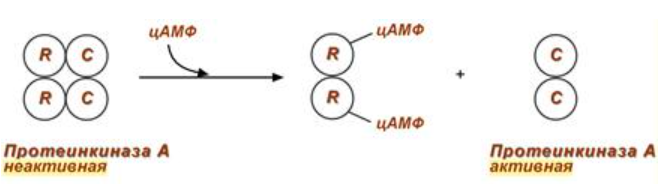
2) Механизм ассоциации-диссоциации:
Регуляция протеинкиназы А через этот механизм
3) Химическая модификация:
- присоединение фосфорной кислоты к белку осуществляют ферменты протеинкиназы, отщепление – протеинфосфатазы
-ферменты могут быть активны как в фосфорилированном, так и в дефосфорилированном состоянии
Пример:
В мышцах ферменты гликогенфосфорилаза и гликогенсинтаза:
При нагрузке: оба фосфорилируются, но при этом синтаза неактивна, а фосфорилиза (на расщепление гликогена работает)
Отдых: оба фермента дефосфорилируются, синтаза при этом становится активной, фосфорилаза – неактивной
4) Частичный протеолиз:
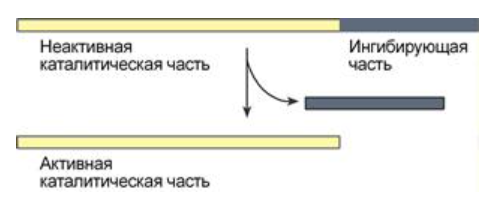
Пример: трипсиноген, пепсиноген