Биохимия 1 коллоквиум
1. Функции белков в организме человека. Физиологически активные пептиды. Уровни структурной организации белков. Первичная, вторичная, третичная и четвертичная структуры белка. Связи, их стабилизирующие. Глобулярные, фибриллярные, трансмембранные белки: особенности строения, примеры. Фолдинг белков. Шапероны. Денатурация и ренативация белков. Прионы и прионные болезни.
Функции белков:
1) Структурная (межклеточное пр-во - протеогликаны, построение органелл - рибосомы)
2) Ферментативная (все ферменты - белки)
3) Гормональная (инсулин, глюкагон - белки, гормоны гипофиза - пептиды)
4) Рецепторная (избирательное связывание гормонов, биологически активных веществ и медиаторов)
5) Транспортная - липопротеины (перенос жира), трансферрин (транспорт железа), Na-K-АТФаза
6) Резервная - белки мышц, эпителиальных тканей, печени - мобилизуются при длительном голодании.
7) Питательная - белки грудного молока, тканевые белки при голодании
8) Защитная - Ig крови, факторы системы комплемента
Физиологически активные пептиды:
1) Нейропептиды - эндорфины, энкефалины
2) Гипоталамические нефрогормоны - либерин и статин видимо
3) Периферические пептиды - брадикинин, ангиотензины,
Уровни структурной организации белков:
Если АМК меньше 10 - пептид;
если от 10 до 40 – полипептид,
если более 40 – белок.
Первичная структура - последовательность АМК
- задается ДНК (при мутациях - другая АМК - другой белок)
- соединены пептидными связями
Вторичная структура - водородные связи между пептидными группами
Атом кислорода одной группы реагирует с атомом водорода следующей, одновременно кислород этой пептидной группы связывается с водородом еще одной
Два возможных варианта вторичной структуры:
- альфа - спираль - правозакрученная, образуется при помощи водородных связей между пептидными группами 1-го и 4-го, 4-го и 7-го, 7-го и 10-го и так далее аминокислотных остатков.
- бета - складчатый слой - белковая молекула лежит "змейкой", удаленные отрезки цепи оказываются рядом и соединяются водородными связями
Обычно присутствуют оба типа, но в глобулярных белках преобладает альфа-спираль, в фибриллярных – бета - складчатый слой
Третичная структура - укладка цепи в глобулу ("клубок").
Четкой границы между 2 и 3 структурами нет в ее основе лежат стерические взаимосвязи между АМК, стоящими далеко друг от друга.
Связи:
- водородные – между ОН-, СООН-, NH2-группами радикалов аминокислот,
- дисульфидные – между остатками цистеина,
- гидрофобные – между остатками алифатических и ароматических аминокислот,
- ионные – между СООН-группами глутамата и аспартата и NH2-группами лизина и аргинина,
- псевдопептидные – между дополнительными СООН-группами глутамата и аспартата и дополнительными NH2-группами лизина и аргинина.
Четвертичная структура - если белки состоят из двух и более полипептидных цепей, связанных между собой нековалентными (не пептидными и не дисульфидными) связями
Стабилизируются водородными связями, ионными и электростатическими взаимодействиями между остатками АМК на поверхности глобулы.
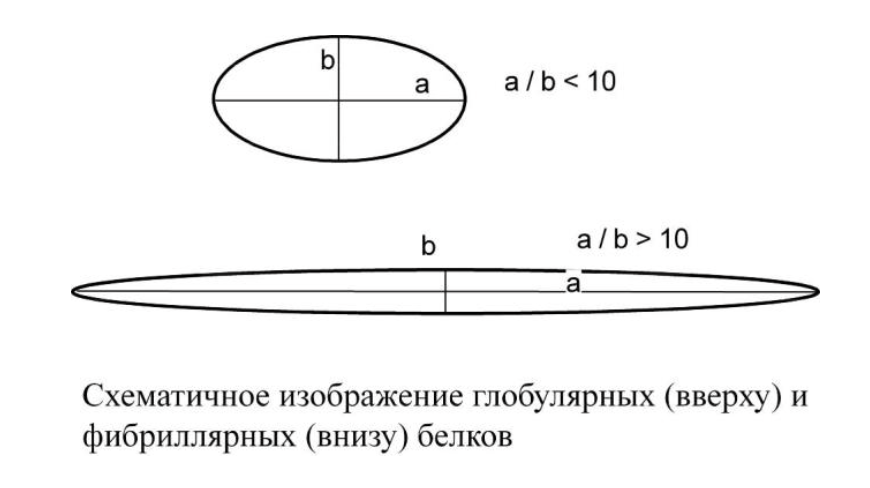
Белки по форме бывают:
Глобулярные – соотношение продольной и поперечной осей составляет <10 (в основном не более 3-4). Они характеризуются компактной укладкой полипептидных цепей. Например, инсулин, альбумин, глобулины плазмы крови.
Фибриллярные – соотношение осей >10. Они состоят из пучков полипептидных цепей, спиралью навитых друг на друга и связанные между собой поперечными ковалентными и водородными связями. Выполняют защитную и структурную функции. Например, кератин, миозин, коллаген.
Трансмембранный белок — мембранный белок, который насквозь пронизывает липидный бислой, в котором он постоянно находится. Плотно закрепляются в мембране при помощи специального класса липидов, называемых кольцевая липидная оболочка. Например, АТФ-синтаза.
ФОЛДИНГ БЕЛКОВ
Фолдинг – процесс укладки полипептидной цепи в правильную трехмерную структуру.
Для фолдинга используется группа вспомогательных белков - шаперонов. Они предотвращают взаимодействие новосинтезированных белков друг с другом, изолируют гидрофобные участки белков от цитоплазмы и "убирают" их внутрь молекулы, правильно располагают белковые домены. В целом шапероны способствуют переходу структуры белков от первичного уровня до третичного и четвертичного.
При нарушении функции шаперонов и отсутствии фолдинга в клетке формируются белковые отложения – развивается амилоидоз.
Дентатурация - необратимое осаждение белка из-за разрыва связей, стабилизирующих 4, 3, 2 структуры белка.
Сопровождается изменением растворимости, вязкости, химической активности, снижением или полной потерей биологической функции.
1. Физическая – температура, УФ, механические воздействия
2. Химическая - кислоты, щелочи, алколоиды, тяжелые металлы
Обратимость осаждения белков обусловлена сохранением первичной структуры белка. Восстановление физико-химических и биологических свойств белка называется ренативация (ренатурация). Иногда для ренативации достаточно просто удалить денатурирующий объект.
ПРИОНЫ И БОЛЕЗНИ
Прионы - инфекционные белки с неправильной третичной структурой. Похожи на вирусы, потому что могут увеличивать свое количество используя функции живых клеток.
Прионы способны катализировать превращение нормальных белков в прионы. Обычно заменяется альфа-спираль на бета-складчатый слой. В итоге получается цепня реакция и накопление.
Прионные болезни:
1) Фатальная семейная бессонница
2) Болезнь Крейтцфельдта — Якоба
Попадают в организм -> катализируют превращение нормальных в прионные -> накапливаются на клетке -> вызывают ее апоптоз -> она для освобождения мембраны синтезирует активные формы кислорода -> формируется очаг воспаления и деструкции
2. Сложные белки. Классификация, строение, примеры. Строение и функции миоглобина
Строение:
У сложных белков, как раз таки кроме белковой цепи (из амк) имеется небелковая часть - лиганд, связанный с белковой цепью. Если лиганд имеет структурную или функциональную нагрузку (или вообще и то и другое сразу) то он переименовывается в простетическую группу.
Классификация:
- нуклеопротеины - белки с НК. Например: рибосомы, хроматин, вирусы. В хроматине - НК связана с белками: гистоновыми (отвечают за начальную упаковку) и негистоновыми.
- фосфопротеины - белки с фосфатной группой. Она связывается с пептидной цепью через остатки тирозина, серина и треонина (тех аминокислот, которые содержат ОН-группу). Например, ферменты: гликогенсинтаза и гликогенфосфорилаза.
На всякий: в клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную (неактивную и активную)
- металлопротеины - с катионами металлов, катион связан с функциональными группами (-COOH или -NH2) амк белка. Примеры: ферритин(запас железа), альфа-амилаза.
- липопротеины - белок с простетической группой в виде липида. В составе могут быть: свободные жирные кислоты, нейтральные жиры, фосфолипиды, холестериды. Хуй знает нужно будет гандонам или нет, но:
Различают четыре основных класса лиипопротеинов:
-липопротеины высокой плотности (ЛПВП, α-липопротеины, α-ЛП),
-липопротеины низкой плотности (ЛПНП, β-липопротеины, β-ЛП),
-липопротеины очень низкой плотности (ЛПОНП)
-хиломикроны (ХМ)
- хромопротеины - с окрашенными простетическими группами. Например: гемопротеины (простетическая группа - гем), ретинальпротеины (простетическая группа - витамин А), флавопротеины (простетическая группа - витамин В2), кобамидпротеины (простетическая группа - витамин В12)
- гликопротеины - содержат углеводный компонент. А вообще, выделяют 2 подкласса белков с углеводами: гликопротеины и протеогликаны. Основное отличие в том, что у гликопротеинов: 15-20% углеводов в составе, у протеогликанов: 80-85% (по названию походу в конце стоит то, чего больше)
Миоглобин:Строение:
Молекула миоглобина образована глобулярным белком с единичной полипептидной цепью и железосодержащим гемом, молекула сходна по строению и функциям с гемоглобином крови.
Функции:
Депо кислорода в мышцах, использование только при сильном уменьшении рО2. Т.о., миоглобин остается оксигенированным до того момента, пока количество кислорода в клетке не снизится до предельно малых величин.
3. Строение и функции гемоглобина. T- и R-формы гемоглобина. Аллостерические эффекты гемоглобина: кооперативный эффект, эффект Бора, эффект 2,3-дифосфоглицерата. Механизмы, физиологическое значение. Особенности фетального гемоглобина. Понятие о гемоглобинопатиях. Серповидно-клеточная анемия. Талассемия.
Строение и функции гемоглобина.
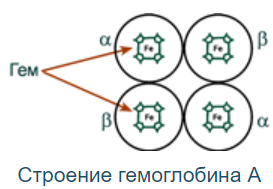
Строение:
Гемоглобин - белок с 4 субъединицами, содержащими гем (картинка). Между собой протомеры соединены связями:
- гидрофобными
- ионными
- водородными
Соединены контактными поверхностями, в десятках точек по принципу комплементарности (разноимённо заряженные группы, гидрофобные участки, неровности на поверхности белка).
T- и R-формы гемоглобина:
Дезоксиформа гемоглобина обозначается как Т-форма, напряженная, она обладает существенно более низким сродством к кислороду. Оксигенированная форма, или R-форма обладает высоким сродством к кислороду.
Кооперативный эффект:
Связывание О2 с одной из субъединиц T-формы приводит к локальным конформационным изменениям, которые ослабляют связь между субъединицами, облегчая присоединение последующих молекул кислорода и изменение других протомеров.
Эффект Бора:
Влияние рН на сродство гемоглобина к кислороду носит название эффекта Бора. При закислении среды сродство снижается, при защелачивании – повышается.
- При повышении концентрации протонов (закисление среды) в тканях возрастает освобождение кислорода из оксигемоглобина.
- В легких после удаления угольной кислоты (в виде СО2) из крови и одновременном увеличении концентрации кислорода высвобождаются ионы Н+ из гемоглобина
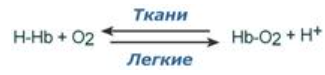
Изменение сродства гемоглобина к кислороду в тканях и в легких (при изменении концентрации ионов H+ и О2) обусловлено конформационными перестройками глобиновой части молекулы.
В ткани:
Молекула О2 отрывается от железа и ионы водорода присоединяются к остаткам гистидина, образуется H-Hb с низким сродством к кислороду.
В легких:
Поступающий в больших количествах кислород "вытесняет" ион водорода из связи с остатком гистидина гемоглобиновой молекулы.
Роль 2,3-дифосфоглицерата
2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта.
2,3-Дифосфоглицерат располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями, образуя поперечный солевой мостик между атомами кислорода 2,3-дифосфоглицератаи аминогруппами обеих β-цепей
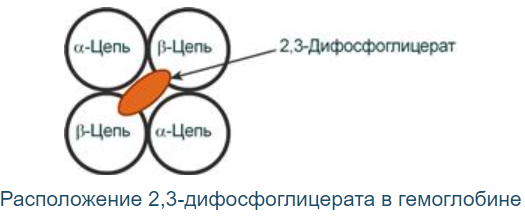
Функция:
Снижает сродство к гемоглобину (то есть отдача О2 в тканях возрастает в 2 раза). Имеет особенное значение при подъеме на высоту и при нехватке кислорода во вдыхаемом воздухе.
Особенности фетального гемоглобина:
- Состоит из 2х альфа и 2х гамма цепей (обычный взрослый из 2х альфа и 2х бета)
- Гемоглобин F обладает повышенным сродством к кислороду и позволяет сравнительно малому объёму крови плода выполнять кислородоснабжающие функции более эффективно
- гемоглобин F обладает меньшей стойкостью к разрушению и меньшей стабильностью в физиологически широком интервале pH и температур
Серповидно-клеточная анемия:
HbS – гемоглобин при серповидно-клеточной анемии. Возникает точковая мутация и в ДНК ЦТТ -> ЦАТ => в бета цепи появляется валин вместо глутамата. Из-за этого поверхность гемоглобина изменяется и появляется "участок прикрепления", за который могут зацепиться другие серповидно-клеточные гемоглобины, поэтому вся эта хуйня соединяется, полимеризуется, происходит деформация эритроцитов и при прохождении через капилляры - гемолиз.
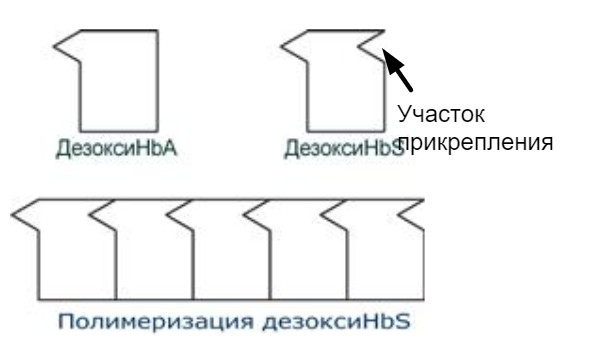
Талассемия:
Для талассемий характерно снижение синтеза α-цепей гемоглобина (α-талассемия) или β-цепей (β-талассемия). Это приводит к нарушению эритропоэза, гемолизу и тяжелым анемиям.
4. Общая характеристика ферментов. Специфичность ферментов. Виды специфичности, примеры. Строение ферментов. Кофакторы и коферменты. Классификация и номенклатура ферментов.
Общая характеристика ферментов
В нормальных физиологических условиях биохимические реакции в организме протекают с высокими скоростями, что обеспечивается биологическими катализаторами белковой природы – ферментами.
Ферменты:
- Катализируют только энергетически возможные реакции
- Не изменяют направления реакции
- Не расходуются в процессе реакции
- Высокая скорость работы
- Высокая специфичность
- Мягкие условия работы (внутри клетки)
- Могут регулировать скорость реакции
- Скорость ферментативной реакции пропорциональна количеству фермента
Специфичность ферментов:
Специфичность - высокая избирательность действия ферментов, основана на комплементарности структуры субстрата и активного центра фермента.
Виды специфичности, примеры:
1) Стереоспецифичность - катализ только одного из стереоизомеров.
Пример: - специфичность ферментов к L-амк
- специфичность к цис- и транс- изомерам (аспартаза реагирует только с транс-изомером – фумаровой кислотой, но не с малеиновой кислотой (цис-изомер))
2) Абсолютная специфичность - главное слово-абсолютная, т.е. один фермент производит катализ только одного вещества.
Пример:- каталаза на Н2О2
- уреаза только мочевины
3) Групповая специфичность - катализ только при наличии определенной связи, хим.группы.
Пример:- наличие пептидной связи для: пепсина, тромбина
- наличие α1,4-гликозидных связей в крахмале и гликогене для: α-амилазы слюнной и поджелудочной желез
- наличие ОН-группы для: алкогольдегидрогеназы
Строение ферментов:
Все ферменты являются белками и обладают всеми свойствами белков. Поэтому, подобно белкам, ферменты делятся на простые и сложные.
Простые:
Только из аминокислот: пепсин, трипсин, лизоцим
Сложные (холоферменты) состоят из 2х частей:
1) Апофермент - белковая часть из амк
2) Кофактор - небелковая часть:
а) Кофермент (НАД+, НАДФ+, ФАД)
б) Простетическая группа (гем, Fe2+, Ca2+, Zn2+)
Примеры:Амилаза (кофактор - Са2+)
Лактатдегидрогеназа (Zn2+)
Сукцинатдегидрогеназа(содержит ФАД)
Классификация и номенклатура ферментов:
I класс – Оксидоредуктазы (ОВР)
название: донор электронов : акцептор электронов – оксидоредуктаза,
пример: алкоголь:НАД-оксидоредуктаза (алкогольдегидрогеназа)
II класс – Трансферазы (перенос различных групп от одного субстрата (донор) к другому (акцептор))
название: донор группы : акцептор группы – переносимая группа трансфераза
пример: АТФ:фруктозо-6-фосфат-фосфотрансфераза (Фосфофруктокиназа)
III класс – Гидролазы (разрыв внутримолекулярных связей в субстрате путем присоединения элементов Н2О)
название: гидролизуемый субстрат : отделяемая группа гидролаза
пример: триацилглицерол : ацилгидролаза (ТАГ-липаза)
IV класс – Лиазы (катализируют разрыв связей без присоединения Н2О)
название: расщепляемый субстрат : отделяемая группа – лиаза
пример: АТФ:дифосфат-лиаза (аденилатциклаза)
V класс – Изомеразы
название: субстрат – [ ] – реакция, где [ ] – обозначение, отражающее суть реакции
пример: α-D-глюкозо-1,6-фосфомутаза (фосфоглюкомутаза)
VI класс – Лигазы (синтез новых веществ)
название: субстрат 1 : субстрат 2 – лигаза
пример: L-глутамат:аммиак-лигаза (Глутаминсинтетаза)
5. Механизм действия ферментов. Активный центр фермента. Этапы ферментативного катализа. Модели взаимодействия фермента с субстратом: «ключ-замок» и Кошланда (индуцированного соответствия).
МЕХАНИЗМ ДЕЙСТВИЯ ФЕРМЕНТОВ
Механизм действия ферментов
Акт катализа складывается из трех последовательных этапов.
1. Образование фермент-субстратного комплекса при взаимодействии через активный центр.
2. Связывание субстрата в нескольких точках активного центра -> изменение структуры субстрата, деформация -> все вместе активация субстрата
Происходит определенная химическая модификация субстрата и превращение его в новый продукт или продукты.
3. В результате такого превращения новое вещество (продукт) утрачивает способность удерживаться в активном центре фермента и фермент-субстратный, вернее уже фермент-продуктный комплекс диссоциирует (распадается).
Виды каталитических реакций:
А+Е = АЕ = БЕ = Е + Б
А+Б+Е = АЕ+Б = АБЕ = АБ + Е
АБ+Е = АБЕ = А+Б+Е, где Е - энзим, А и Б - субстраты, либо продукты реакции.
АКТИВНЫЙ ЦЕНТР ФЕРМЕНТА
Активный центр - комбинация АМК остатков, обеспечивающая связывание субстрата и осуществляющая катализ.
- якорный участок - связывание и ориентация субстрата в активном центре
- каталитический участок - сама реакция
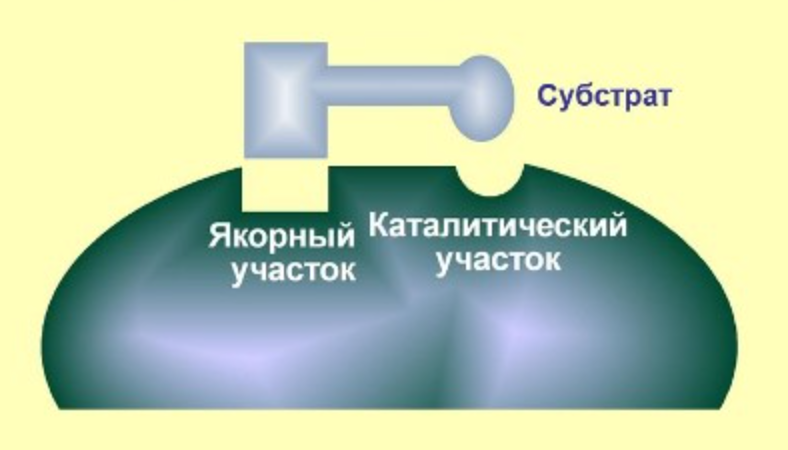
У ферментов, имеющих в своем составе несколько мономеров, может быть несколько активных центров по числу субъединиц. Также две и более субъединицы могут формировать один активный центр.
У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора. Например, в реакции превращения пировиноградной кислоты (пируват) в молочную кислоту (лактат) сначала к апоферменту лактатдегидрогеназы присоединяется НАД, формируется активный центр, и только потом входит пируват
ЭТАПЫ ФЕРМЕНТАТИВНОГО КАТАЛИЗА
1. Присоединение субстрата (S) к ферменту (E) с образованием фермент-субстратного комплекса (E-S).
2. Преобразование фермент-субстратного комплекса в один или несколько переходных комплексов (E-X) за одну или несколько стадий.
3. Превращение переходного комплекса в комплекс фермент-продукт (E-P).
4. Отделение конечных продуктов от фермента.
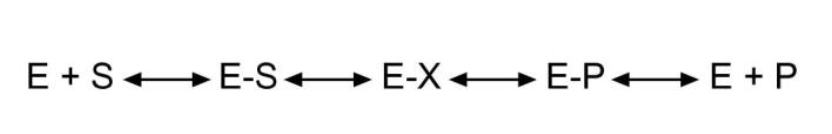
КЛЮЧ-ЗАМОК
Гипотеза Фишера "ключ-замок") – активный центр фермента строго соответствует конфигурации субстрата и не изменяется при его присоединении. Эта модель хорошо объясняет абсолютную специфичность, но не групповую.
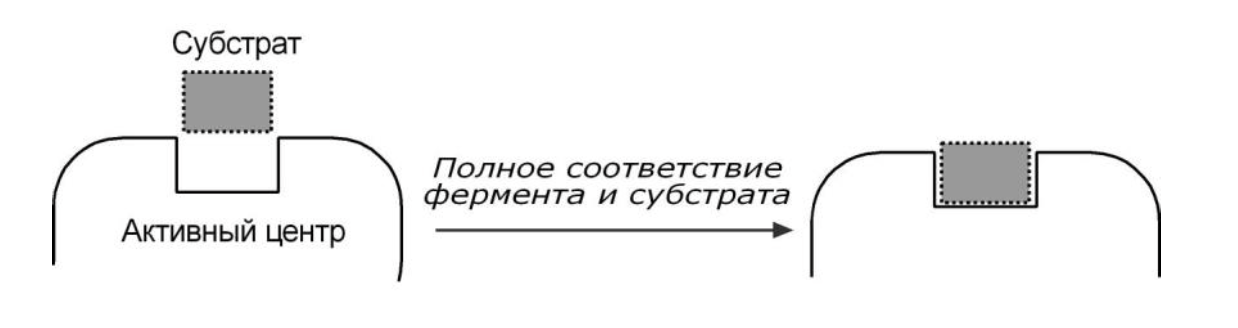
ГИПОТЕЗА КОШЛАНДА
Гипотеза Кошланда (модель "индуцированного соответствия") – подразумевает гибкость активного центра. Присоединение субстрата к якорному участку фермента вызывает изменение конфигурации каталитического центра таким образом, чтобы его форма соответствовала форме субстрата.

6. Основы кинетики ферментативных реакций. Зависимость скорости ферментативной реакции от количества субстрата. Уравнение Михаэлиса- Ментен. Зависимость скорости ферментативной реакции от количества фермента. Зависимость скорости ферментативной реакции от температуры и рН среды.
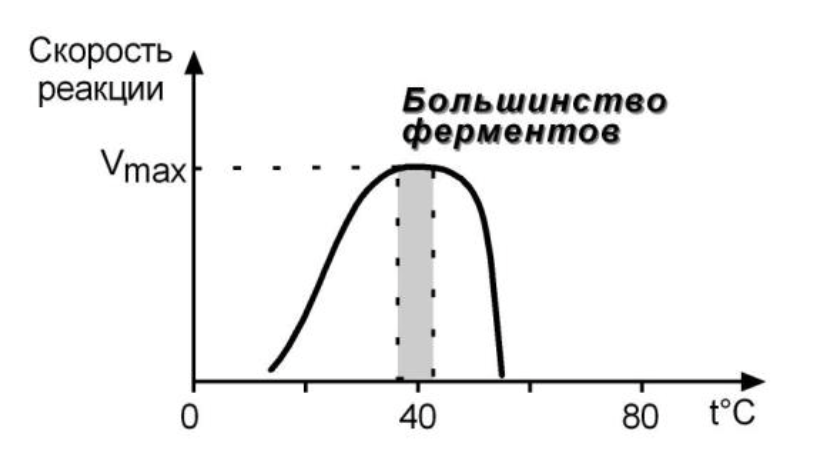
ЗАВИСИМОСТЬ ОТ ТЕМПЕРАТУРЫ
Колоколообразная кривая с оптимумом для каждого фермента.
Чем выше t - тем больше скорость, но только в пределах 55-60 градусов (денатурация)
но есть микроорганизмы в гейзерах
При понижении t - скорость снижается, но не исчезает (спячка ежей)
ЗАВИСИМОСТЬ ОТ pH
Колоколообразная кривая с оптимумом для каждого фермента.
У каждого фермента узкий интервал активности (пепсин 1,5-2,5,
трипсин 8,0-8,5, амилаза слюны 7,2,),
Так как
изменение pH -> будет меняться заряд АМК -> лизин, аргинин, глутамат, аспартат -> изменение конформации -> не может связаться с субстратом
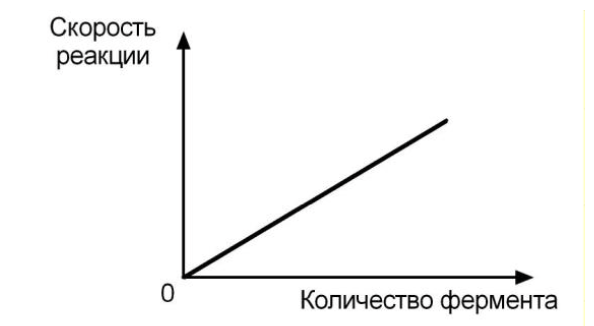
ЗАВИСИМОСТЬ ОТ КОЛИЧЕСТВА ФЕРМЕНТА
При увеличении количества молекул фермента скорость реакции возрастает непрерывно и прямо пропорционально количеству фермента, т.к. большее количество молекул фермента производит большее число молекул продукта
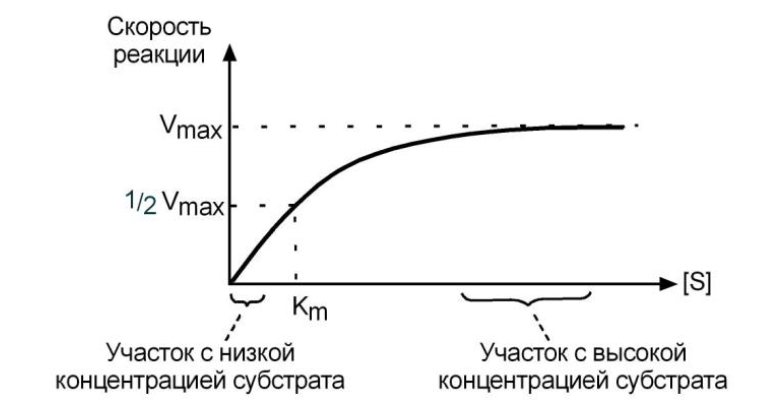
ЗАВИСИМОСТЬ ОТ КОЛИЧЕСТВА СУБСТРАТА
При увеличении концентрации субстрата
1) скорость сначала возрастает, т.к. подключаются новые и новые молекулы фермента.
Т.е. скорость накопления продукта возрастает, и это означает увеличение активности фермента.
2) Затем наблюдается эффект насыщения (плато на кривой), когда все молекулы фермента заняты молекулами субстрата и непрерывно ведут катализ - скорость реакции максимальна.
3) В некоторых случаях, при дальнейшем увеличении концентрации субстрата между его молекулами возникает конкуренция за активный центр фермента и активность фермента (скорость реакции) снижается.
УРАВНЕНИЕ МИХАЭЛИСА-МЕНТЕН
Зависимость активности фермента от субстрата описывает уравнение Михаэлиса- Ментен, использующее константу Михаэлиса (Km)
Биологический смысл - увеличение величины Кm означает снижение сродства фермента к субстрату.
Уравнение Михаэлиса-Ментен показывает взаимосвязь максимальной и реальной скоростей реакции, константы Михаэлиса и концентрации субстрата.
В математическом смысле Km соответствует концентрации субстрата при которой скорость реакции равна половине максимальной.
Выделяют три решения уравнения:
1. Концентрация субстрата равна величине констансты Михаэлиса ([S] = Km). В этом случае скорость реакции V будет равна половине максимальной Vmax. (V = 1⁄2 Vmax).
2. Концентрация субстрата [S] значительно больше Km ([S] >> Km). В этом случае, величиной Km можно пренебречь и при решении получаем, что скорость реакции максимальна (плато на графике).
3. Концентрация субстрата значительно меньше Km ([S] << Km). В этом случае, знаме- натель уравнения мало изменяется при изменении [S], а величина скорости реакции V прямо пропорциональна [S] (график линеен).
7. Ингибирование ферментативной активности. Конкурентное и неконкурентное обратимое ингибирование. Необратимое ингибирование. Кинетические зависимости. Примеры.
Ингибирование ферментативной активности
Два основных пути ингибирования:
а) По прочности связывания: обратимое и необратимое
б) По отношению ингибитора к активному центру: конкурентное и неконкурентное
Конкурентное и неконкурентное обратимое ингибирование
При обратимом ингибировании происходит непрочное связывание ингибитора с функциональными группами фермента, вследствие чего активность постепенно восстанавливается.
Конкурентное ингибирование:
ингибитор похож на субстрат фермента => соперничество за акт.центр фермента => уменьшение связывания субстрата с ферментом и нарушению катализа
-при данном ингибировании максимальная скорость реакции остается вполне достижимой при создании высоких концентраций субстрата
Пример:
- ингибирование сукцинат-дегидрогеназы малонатом, который похож на сукцинат
- антиметаболиты (сульфаниламиды против бактерий)
Неконкурентное ингибирование:
Присоединением ингибитора не в активном центре, а в другом месте молекулы => изменение структуры акт.центра => связь акт.центра с субстратом невозможна
Пример:
- синильная кислота связывается с гемовым железом ферментов дыхательной цепи и блокирует клеточное дыхание
- ионы тяжелых металлов (Cu2+, Hg2+, Ag+) c SH- группами белков
!!!Изменение концентрации субстрата никак не влияет
Необратимое ингибирование:
Связывание или разрушение функциональных групп фермента, необходимых для его активности
Пример:
Диизопропилфторфосфат прочно и необратимо связывается с гидроксигруппой серина в активном центре фермента ацетилхолинэстеразы
8. Аллостерические ферменты. Особенности строения и функционирования. Аллостерические эффекторы. Регуляция активности ферментов белок- белковыми взаимодействиями. Регуляторные белки. Ассоциация- диссоциация. Примеры. Регуляция активности ферментов путем фосфорилирования-дефосфорилирования, частичным протеолизом.
АЛЛОСТЕРИЧЕСКИЕ ФЕРМЕНТЫ
- ферменты, активность которых регулирует не только наличие субстрата но и регуляторные молекулы
У них помимо активного центра есть аллостерический центр.
Аллостерический центр - пространственно отделен от активного центра.
С активным центром связывается активатор или ингибитор -> изменяется конформация белка -> активность фермента
Аллостерический и активный центр находятся в разных субъединицах, поэтому обычно АФ - сложные белки.
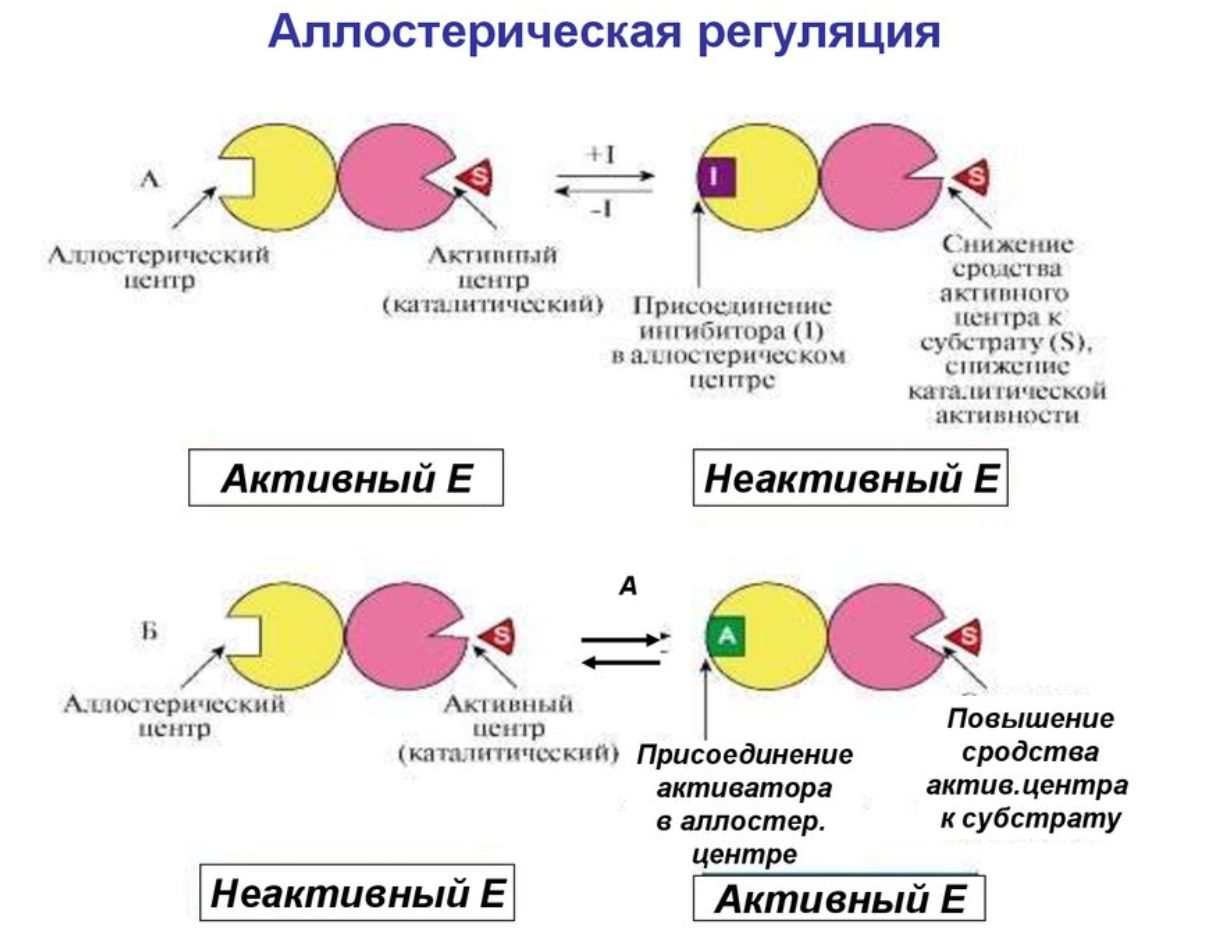
В качестве отрицательного регулятора может выступать конечный или промежуточный метаболит биохимического процесса или продукт данной реакции, т.е включается механизм обратной отрицательной связи.
Если регуляторами являются начальный метаболит или субстрат реакции, то говорят о прямой регуляции, она может быть как положительной, так и отрицательной. Также регулятором могут быть метаболиты биохимических путей, каким то образом связанных с данной реакцией.
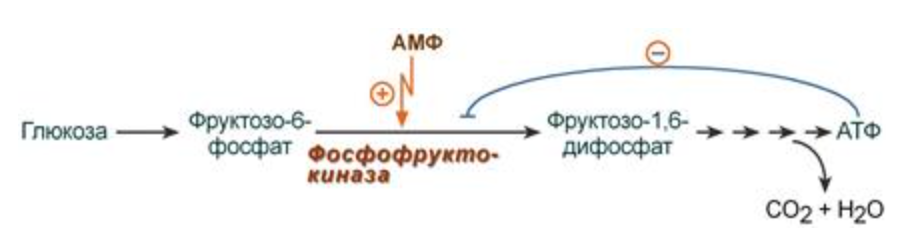
ЭФФЕКТОРЫ
- это и есть активаторы или ингибиторы - ими могут быть:
1) метаболиты и гормоны
2) лекарства
3) яды
В случае с метаболитами:
1) Если конечный продукт - ингибтор, а начальный - активатор - гетеротропная регуляция - очень часто в процессах метаболизма
2) Если субстрат и эффектор - одно и то же в-во - гомотропная регуляция
Регуляторные белки:
- белки-рецепторы, воспринимающие сигнал
- белки-гормоны и другие вещества, передающие сигналы клеткам
Бля речь про белок-белковое взаимодействие: после влияния каких-либо факторов на специфичные белки изменяется активность этих белков, и они, в свою очередь, воздействуют на нужный фермент.
1)
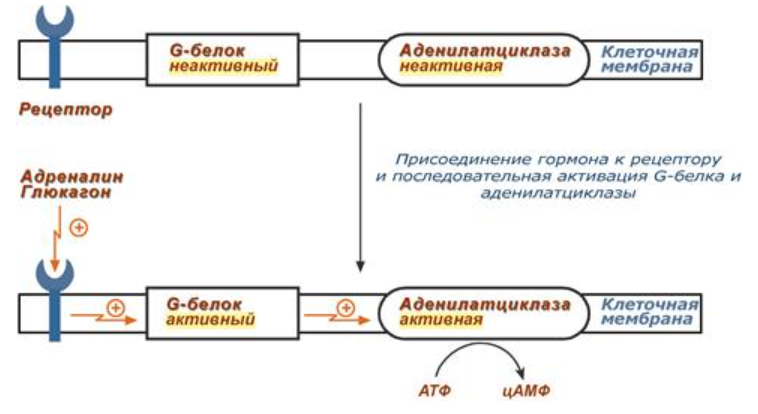
2) Механизм ассоциации-диссоциации:
Регуляция протеинкиназы А через этот механизм
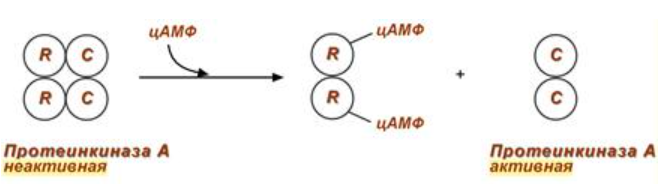
3) Химическая модификация:
- присоединение фосфорной кислоты к белку осуществляют ферменты протеинкиназы, отщепление – протеинфосфатазы
-ферменты могут быть активны как в фосфорилированном, так и в дефосфорилированном состоянии
Пример:
В мышцах ферменты гликогенфосфорилаза и гликогенсинтаза:
При нагрузке: оба фосфорилируются, но при этом синтаза неактивна, а фосфорилиза (на расщепление гликогена работает)
Отдых: оба фермента дефосфорилируются, синтаза при этом становится активной, фосфорилаза – неактивной
4) Частичный протеолиз:
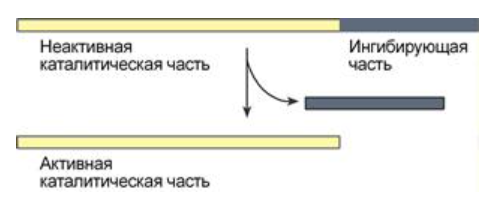
Пример: трипсиноген, пепсиноген
9. Изоферменты. Примеры. Биологическое значение. Понятие об энзимопатиях. Примеры. Энзимодиагностика и энзимотерапия. Примеры использования ферментов в качестве лекарственных средств. Понятие об абзимах.
ИЗОФЕРМЕНТЫ
Изоферменты - это молекулярные формы одного и того же фермента, но с небольшими ген. изменениями в первичной структуре , но катализирующие одну и ту же реакцию.
- катализируют одну реакцию, НО
- разная чувствительность к активаторам и ингибиторам
- разный оптимум работы pH
Как правило, изоферменты имеют четвертичную структуру - состоят из двух или более субъединиц.
Например, димерный фермент креатинкиназа представлен тремя изо- ферментными формами, составленными из двух типов субъединиц: M и B. Креатинкиназа-1 состоит из субъединиц типа B, локализуется в головном мозге, креатинкиназа-2 – по одной М и В субъединице активна в миокарде,
креатинкиназа-3 – две М-субъединицы, специфична для скелетной мышцы.
Биологическая роль: как я поняла - адаптивность реакций, типа так как разная регуляция работы, процесс не останавливается при внешних и внутренних воздействиях
ЭНЗИМОДИАГНОСТИКА
Энзимодиагностика – исследование активности ферментов для диагностики заболеваний.
Примеры:
- лактатдегидрогеназа, ее активности в плазме необходимо при заболеваниях сердца, печени, скелетной мускулатуры, увеличение при инфаркте, потому что выливается при разрушении кардиомиоцитов
- увеличение активности альфа-амилазы в плазме и моче наблюдается при воспалительных процессах в поджелудочной и слюнных железах.
ЭНЗИМОТЕРАПИЯ
Энзимотерапия – использование ферментов в качестве лекарственных средств.
Чаще всего препараты - комплексы ферментов ЖКТ пепсина, химотрипссина, липазы, амилазы и других (Фестал, Панзинорм форте, Мезим форте, Энзистал и т.п.) для заместительной терапии при нарушениях переваривания в ЖКТ
- Гиалуронидаза - для размягчения рубцов, появления подвижности в суставах, рассасывания гематом
- Цитохром с – при асфиксии новорожденных, астме
- Трипсин - для разжижения густой и вяз- кой мокроты.
ЭНЗИМОПАТИИ
Энзимопатии - состояния, связанные с патологическим изменением активности ферментов. Чаще всего нарушения метаболизма и снижение активности.
Из-за нарушений происходит накопление каких-нибудь субстратов реакции (при фенилкетонурии) или недостаток (альбинизм)
Первичные энзимопатии - генетический дефект.
Например, - фенилкетонурия связана с дефектом фенилаланин-4-монооксигеназы, которая превращает фенилаланин в тирозин - подагра - дефекто ферментов метаболизма пуриновых оснований и накоплением мочевой кислоты
Вторичные энзимопатии - следствие заболеваний органов, вирусных инфекций
-> нарушение синтеза фермента или условий его работы
Например, - гипераммониемия при заболеваниях печени, при которых ухудшается синтез мочевины и в крови накапливается аммиак - недостаточность ферментов ЖКТ при заболеваниях желудка, поджелудочной или желчного
Абзимы
Абзимы - антитела, имеющие каталитическую функцию и катализирующие конкретные реакции.
Такая способность возникает в результате формирования промежуточного продукта при связывании антитела с антигеном