Биохимия Биологическое окисление
1. Строение биологических мембран. Липиды мембран: состав, функции. Белки мембран. Разновидности, функции. Свойства биологических мембран: микровязкость, асимметрия. Жидкостно-мозаичная модель строения мембран.
Все живые клетки отделены от окружающей среды поверхностью называемой клеточной мембраной.
У эукариотов образуют компартменты - мембранные органеллы - ядро, митохондрии.
Мембраны - не просто поверхности, но и все транспортные системы, рецепторы и тд, встроенные в них.
Химический состав
Мембраны состоят из липидных и белковых молекул, относительное количество которых варьирует - от 1/5 — белок до 3/4 — белок у разных мембран.
Углеводы содержатся в форме гликопротеинов, гликолипидов и составляют 0,5—10% вещества мембраны.
Белки мембран
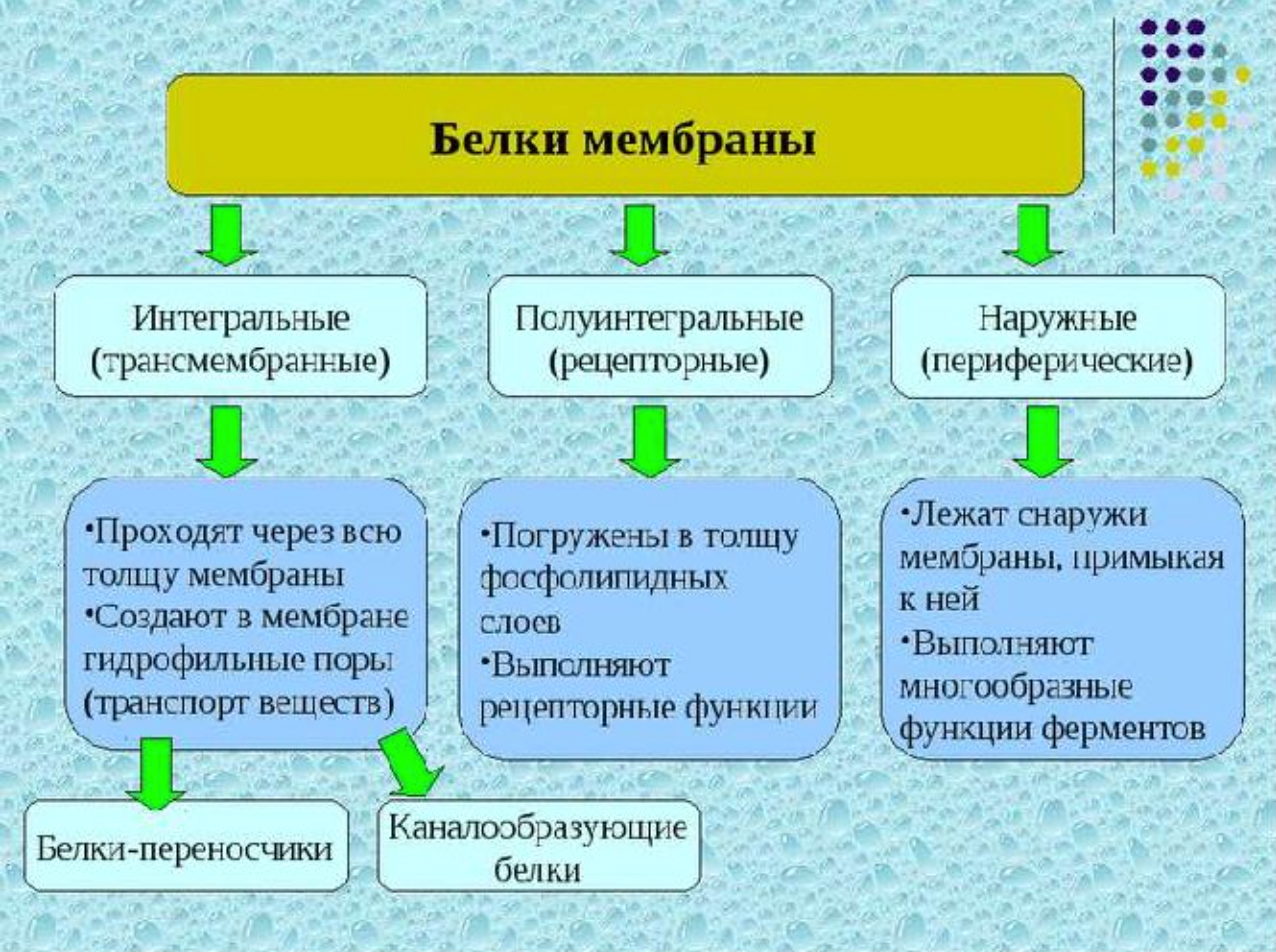
Липиды мембран
Стенки мембран образованы липидами трёх классов:
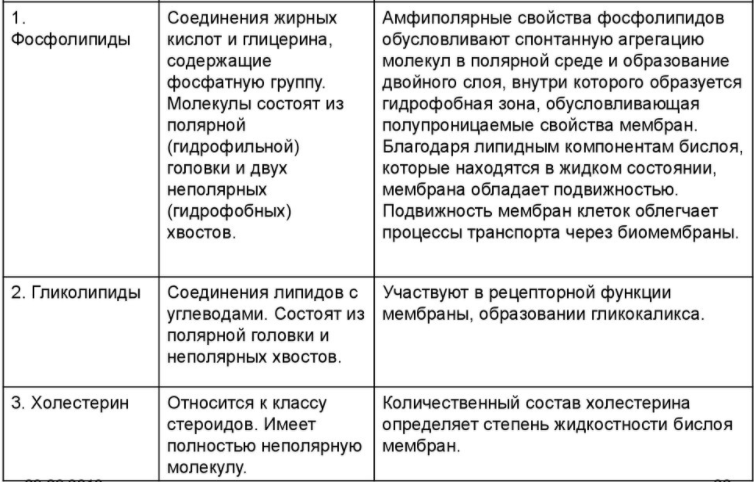
Эти вещества образуют жидкостно-мозаичную структуру, состоящую из трёх слоёв. Фосфолипиды формируют два внешних слоя. Они имеют гидрофильную головку, от которой отходят два гидрофобных хвостика. Хвостики повёрнуты внутрь структуры, образуя внутренний слой. При встраивании холестерола в хвостики фосфолипидов мембрана приобретает жёсткость.
Микровязкость
Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии.
Скорость перемещения молекул зависит от микровязкости мембран, которая, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов.
Преобладают ненасыщенные - микровязкость больше
Преобладают насыщенные - микровязкость меньше
Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые "изломы". Эти "изломы" препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно и более "текучей".
На текучесть мембран также влияют размеры углеводородных "хвостов" липидов, с увеличением длины которых мембрана становится более "текучей".
Ассиметрия

Липидный состав монослоев различен. Например, в плазматической мембране фосфатидилхолин преобладает в наружном слое, а фосфатидилсерин во внутреннем слое. Углеводные части белков и липидов располагаются на наружной части мембраны. Кроме того, поверхности мембраны отличаются по составу белков. Степень такой асимметрии мембран различна у разных типов мембран и может меняться в процессе жизнедеятельности клетки и ее старения.
Жидкостно-мозаичная модель мембраны
Согласно этой модели, мембрана - два параллельных слоя липидов (липидный бислой). У липидов гидрофильная головка (сахар, фосфат..) и гидрофобный хвост (остатки жирных кислот). Хвосты спрятаны внутрь, головки наружу. Хвосты удерживаются вместе гидрофобными взаимодействиями.
То есть по данной модели при физиологических температурах мембрана представляет текучий фосфолипидный бислой, в который погружены свободно-диффундирующие белки.
Молекулы белков не образуют сплошного слоя, а погружены в ФЛ слой на разную глубину в виде мозаики.
Мембраны ассиметричны - разное соотношение белков и липидов во внешнем и внутреннем слое, углеводные компоненты связаны с липидом или белком и их цепи обращены во внеклеточную среду.
2. Транспорт веществ через биологические мембраны. Пассивный транспорт. Ионные каналы. Аквапорины. Активный транспорт веществ через мембраны. Первично-активный транспорт. Строение транспортных АТФ-аз. Вторичноактивный транспорт. Эндоцитоз и экзоцитоз: биохимические механизмы.
Транспорт веществ через биологические мембраны:
Весь транспорт делится на активный и пассивный:
Пассивный:
- Осмос
- Простая диффузия
- Облегченная диффузия
Активный:
- Первично-активный транспорт
- Вторично-активный транспорт
- Везикулярный транспорт: эндоцитоз (пино-, фагоцитоз) и экзоцитоз
Пассивный транспорт:
1) Осмос - движение жидкости, газов из области с меньшей концентрацией в большую
2) Простая диффузия - просто перемещение частиц туда-сюда в следствии их движения (без транспортных белков, просто через билипидный слой)
3) Облегченная диффузия - транспорт молекул через мембрану при помощи специальных транспортных белков
Ионные каналы
Это порообразующие белки, осуществляют облегченную диффузию. Ну тут можно просто шизу воткнуть, что есть независимые, потенциал-зависимые, лиганд-зависимые, механочувствительные, также про работу створок канала
Аквапорины
- Ионные каналы, отвечающие за транспорт воды.
- Избирательно пропускают молекулы воды, позволяя ей поступать в клетку и покидать её, в то же время препятствуя протоку ионов и других растворимых веществ.
- Регулируются фосфорилированием, которое приводит к большей активности аквапорина
Активный транспорт веществ через мембраны. Первично-активный транспорт:
Здесь уже участвуют ионные насосы, а не каналы.
Насосы переносят вещества против градиента концентрации за счет энергии гидролиза АТФ
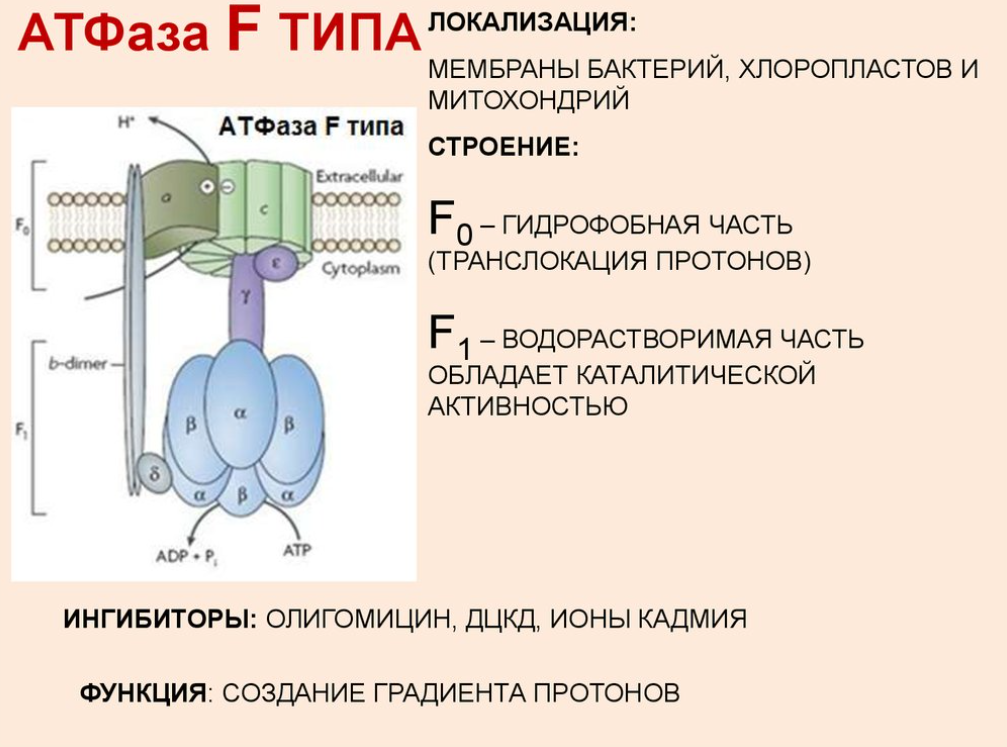
Строение транспортных АТФ-аз:
АТФазы - ферменты, гидролазы. Отсоединяют от АТФ 1 или 2 остатка фосфорной кислоты с высвобождением энергии.
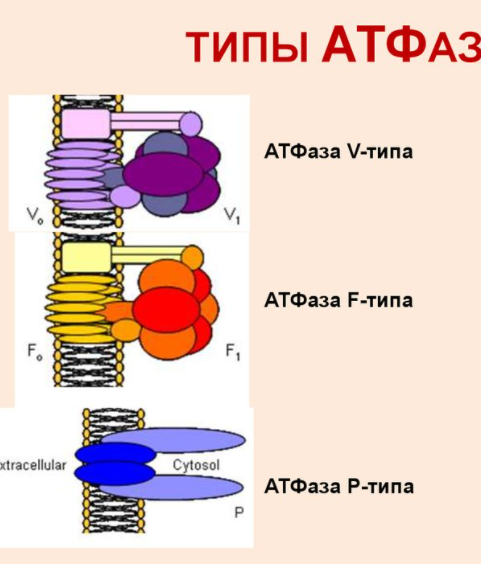
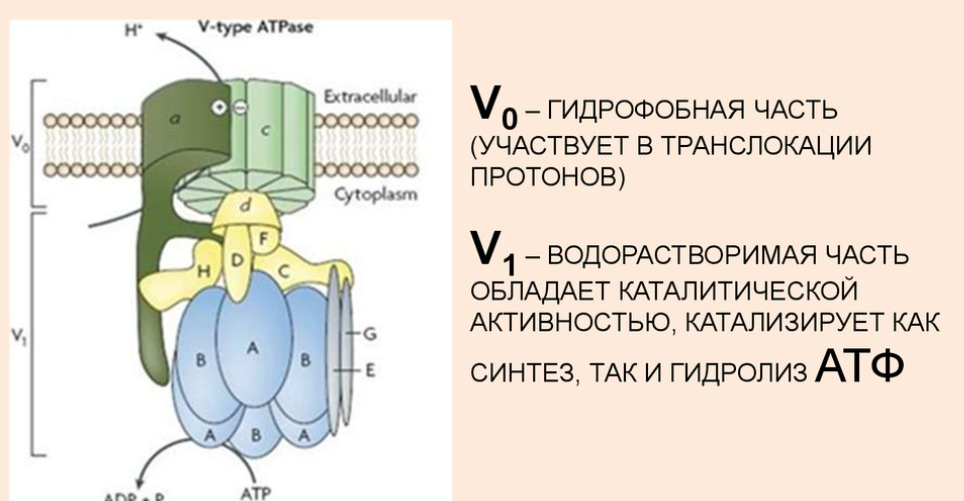
Переносят протоны

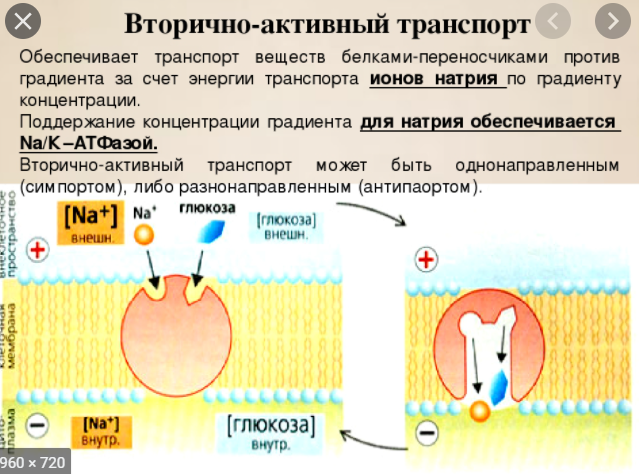
Вторичноактивный транспорт.
Эндоцитоз и экзоцитоз: биохимические механизмы
Эндоцитоз - перенос веществ из среды в клетку вместе с частью плазматической мембраны, путем образования мембраной пузырьков.
Обычно клетки формируют пузырьки постоянно, ритмично и постоянно что-то захватывают из внеклеточной жидкости, но иногда эндоцитоз индуцируется веществами, контактирующими с клеткой. Например, рецепто ры плазматической мембраны гепатоцитов улавливают многие гликопротеины плазмы крови, а затем эти белки эндоцитируются.
Эндоцитоз: - процесс, нуждающийся в энергии
- источником последней служит АТФ
- потребителем энергии являются сокращающиеся микрофиламенты
- присоединение лиганда к рецепторам изменяет их конформацию => после чего к ним присоединяются микрофибриллы => их сокращение приводит к впячиванию мембраны и отделению пузырька
Экзоцитоз:
Есть два основных типа экзоцитоза в эукариот: регулируемые и нерегулируемые (или составные). Регулируемый экзоцитоз, который требует внешнего сигнала, используется для высвобождения нейротрансмиттеров и секреции гормонов. В отличие от регулируемого экзоцитоза, составной экзоцитоз осуществляется всеми клетками. Клетки используют составной экзоцитоз для высвобождения компонентов внеклеточной матрицы или включения белков в плазменную мембрану.
Моторные белки активно перемещают пузырьки по цитоскелетам микротрубочек и нитей. Вторым шагом является привязка пузырьков, в которой пузырьки связываются с плазменной мембраной. На третьем этапе, стыковке пузырьков, мембрана пузырьков прикрепляется к плазменной мембране, и две мембраны начинают сливаться.
Четвертый шаг, праймирование пузырьков, происходит только при регулируемом экзоцитозе. Праймирование включает в себя изменения, происходящие после привязки пузырька, но прежде чем он выпускает свое содержимое. Праймирование готовит пузырьки для синтеза с плазменной мембраной.
Пятый шаг - слияние пузырьков. Оно может быть полным или по схеме "поцеловал-и-убежал". При полном слиянии пузырьки полностью разрушаются и становятся частью плазменной мембраны, изгоняя их содержимое из клетки в процессе.
3. Трансмембранная передача сигнала. Белки-рецепторы. G-белки: строение, функции, регуляция.
Важное свойство мембран - способность воспринимать и передавать внутрь клетки сигналы из внешней среды. "Узнавание" сигнальных молекул осуществляется с помощью белков-рецепторов, встроенных в клеточную мембрану клеток-мишеней или находящихся в клетке. Клетку-мишень определяют по способности избирательно связывать данную сигнальную молекулу с помощью рецептора.
Белки - рецепторы
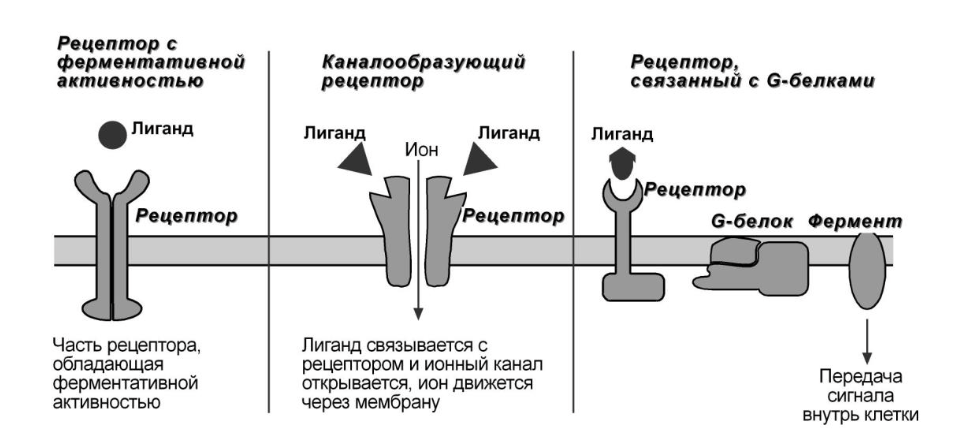
1. Каналообразующие рецепторы – присоединение лиганда к рецептору вызывает открытие ионного канала на мембране.
Нейромедиаторы (ацетилхолин, глицин, ГАМК, серотонин)
2. Рецепторы, обладающие каталитической активностью – при взаимодействии лиганда с рецептором активируется внутриклеточный домен рецептора, имеющий ферментативную активность (тиронинкиназа, тирозинфосфатаза, гуанилатцикла- за).
Рецепторы к цитокинам (интерфероны, интерлейкины, факторы роста), к инсулину, пролактину и СТГ.
3. Рецепторы, связанные с G-белками – передача сигнала от гормона происходит при посредстве G-белка. G-белок влияет на ферменты, образующие вторичные посредники (мессенджеры).
Последние передают сигнал на внутриклеточные белки.
Аденилатциклазный и кальций-фосфолипидный механизмы. Через такие рецепторы действуют часть медиаторов и пептидные гормоны
G-белки
Строение: Они состоят из трёх субъединиц- альфа, бета и гамма.
Бета и гамма - всегда вместе, когда лиганд связывается с рецептором, диссоциируют вместе, поэтому их называют бета-гамма-комплекс или бета-гамма-димер.
Функции: передача сигнала
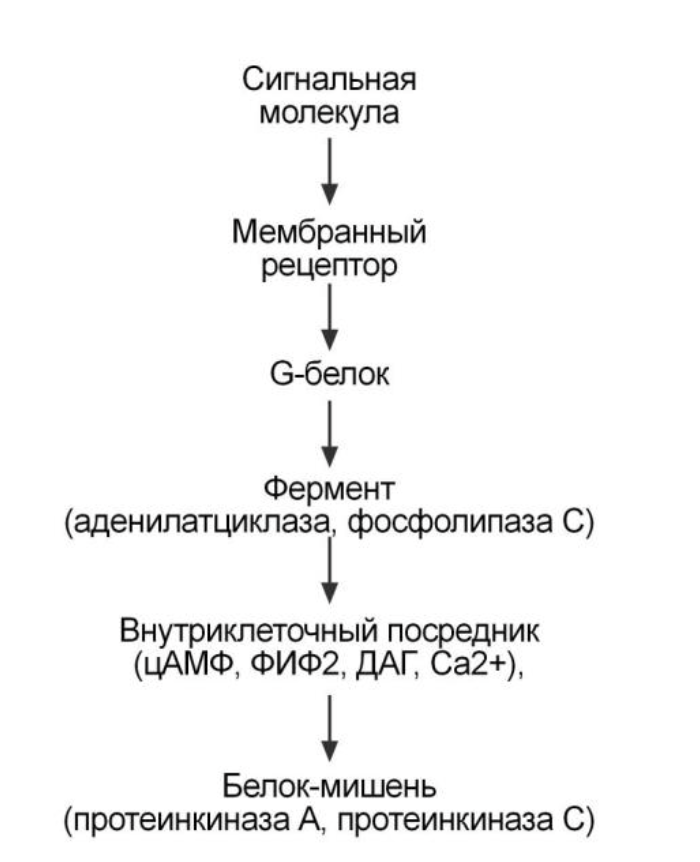
Аденилатциклазный механизм
1. G-белок, который состоит из трех субъединиц, альфа-субъединица связана с ГДФ.
В составе G-белков α-субъединицы бывают двух типов по отношению к ферменту: активирующие αs и ингибирующие αi.
2. В результате взаимодействия с рецептором бета и гамма-субъединицы отщепляются, одновременно в αS-субъединице ГДФ заменяется на ГТФ.
3. Активированная таким образом αS-субъединица стимулирует аденилатциклазу, которая начинает синтез цАМФ.
и дальше...
4. Катаболизм и анаболизм. Общая схема обмена веществ и энергии в организме человека. Специфические и общие пути катаболизма.
Анаболизм
Анаболизм – это биосинтез белков, полисахаридов, липидов, нуклеиновых кислот и других макромолекул из малых молекул-предшественников. Поскольку он сопровождается усложнением структуры, то требует затрат энергии. Источником такой энергии является энергия АТФ.
Катаболизм
Катаболизм – расщепление и окисление сложных органических молекул до более простых конечных продуктов. Оно сопровождается высвобождением энергии, заключенной в сложной структуре веществ. Большая часть высвобожденной энергии рассеивается в виде тепла. Меньшая часть этой энергии "перехватывается" коферментами окислительных реакций НАД и ФАД, некоторая часть сразу используется для синтеза АТФ.
Общая схема обмена веществ и энергии в организме человека:
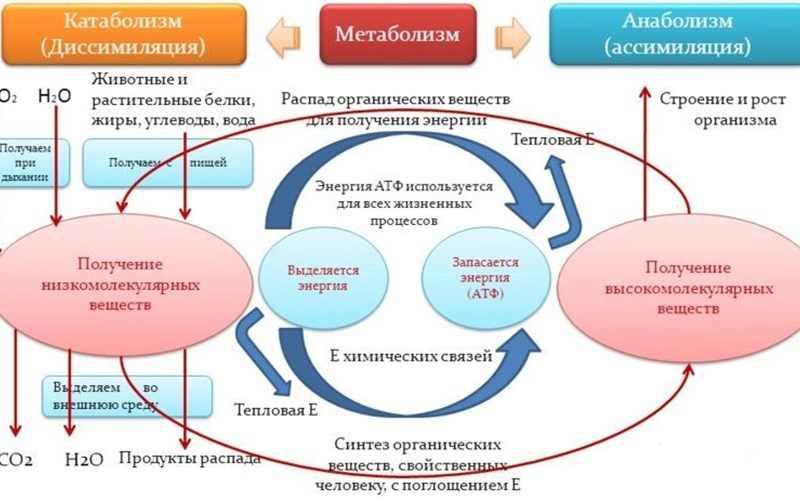
Специфические и общие пути катаболизма:
В катаболизме различают три стадии:
1) Полимеры превращаются в мономеры (белки – в аминокислоты, углеводы в моносахариды, липиды – в глицерол и жирные кислоты). Химическая энергия при этом рассеивается в виде тепла.
2) Мономеры превращаются в общие продукты, в подавляющем большинстве в ацетил-КоА. Химическая энергия частично рассеивается в виде тепла, частично накапливается в виде восстановленных коферментных форм (НАДН, ФАДН2), частично запасается в макроэргических связях АТФ (субстратное фосфорилирование).
1-ая и 2-ая стадии катаболизма относятся к специфическим путям, которые уникальны для метаболизма белков, липидов и углеводов.
3) Заключительный этап катаболизма, сводится к окислению ацетил-КоА до СО2 и Н2О в реакциях цикла трикарбоновых кислот (цикла Кребса) – общий путь катаболизма. Окислительные реакции общего пути катаболизма сопряжены с цепью тканевого дыхания. При этом энергия (40-45%) запасается в виде АТФ (окислительное фосфорилирование).
5. Окислительное декарбоксилирование пирувата. Строение пируватдегидрогеназного комплекса. Стадии окислительного декарбоксилирования пирувата. Регуляция пируватдегидрогеназного комплекса.
Смысл комплекса: Декарбоксилируется пируват, восстановливается НАД до НАДН и образуется ацетил-SKoA.
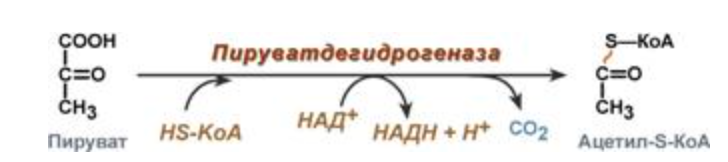
Строение
Превращение состоит из пяти последовательных реакций, осуществляется мультиферментным комплексом, прикрепленным к внутренней митохондриальной мембране со стороны матрикса. В составе комплекса насчитывают 3 фермента и 5 коферментов:
- Пируватдегидрогеназа (Е1, ПВК-дегидрогеназа), ее коферментом является тиаминдифосфат (ТДФ), катализирует 1-ю реакцию.
- Дигидролипоат-ацетилтрансфераза (Е2), ее коферментом является липоевая кислота, катализирует 2-ю и 3-ю реакции.
- Дигидролипоат-дегидрогеназа (Е3), кофермент – ФАД, катализирует 4-ю и 5-ю реакции.
Помимо указанных коферментов, которые прочно связаны с соответствующими ферментами, в работе комплекса принимают участие коэнзим А и НАД.
Суть первых трех реакций декарбоксилирование пирувата, его окисление до ацетила и перенос ацетила на коэнзим А.
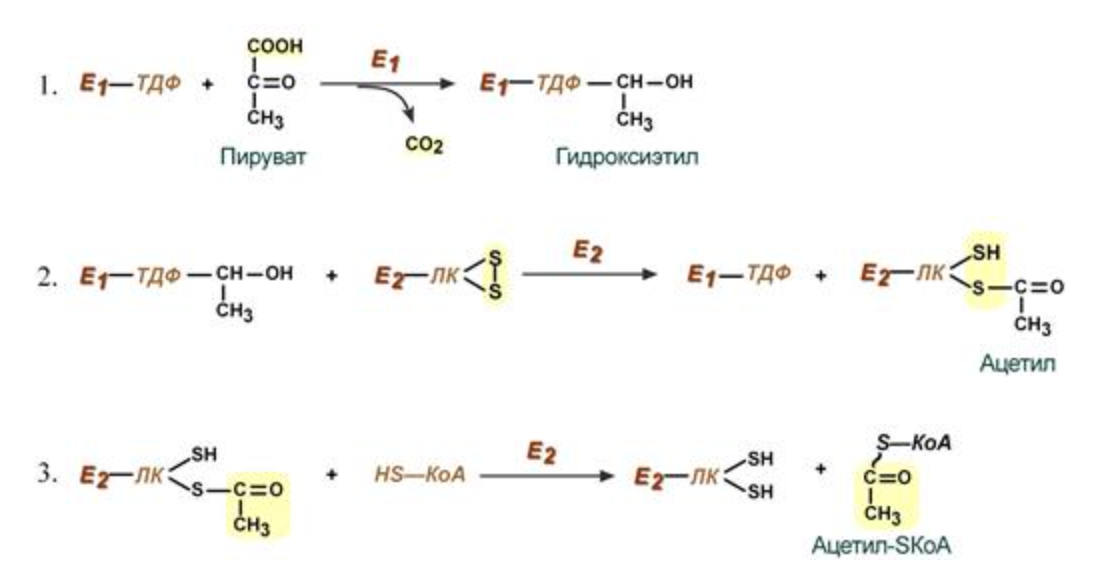
1 реакция пируват -> гидроксиэтил (Пируватдегидрогеназа, кофермент - тиаминдифосфат)
2 реакция гидроксиэтил -> ацетил (дигидролипоат-ацетилтрансфераза, кофермент - липоевая кислота)
3 реакция ацетил -> ацетил-SKoa (дигидролипоат-ацетилтрансфераза, кофермент - липоевая кислота)
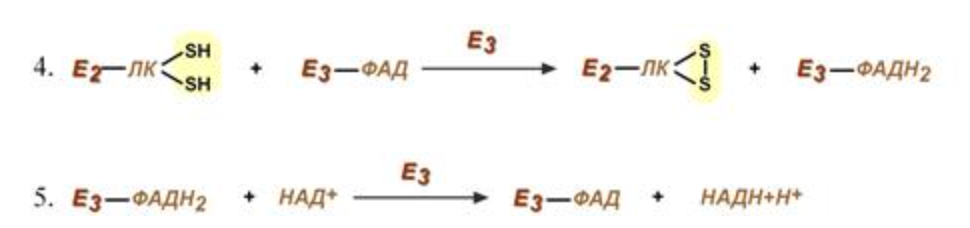
4 и 5 реакции Липоевая кислота и ФАД - в окисленное состояние - образуется НАДН (катализируются дигидролипоат-дегидрогеназой, кофермент - ФАД)
Регуляция
2 вспомогательных фермента – киназа и фосфатаза, регулируют активность пируватдегидрогеназы путем фосфорилирования и дефосфорилирования.
Киназа активируется при избытке АТФ и продуктов реакции, фосфорилирует пируватдегидрогеназу, инактивирует.
Фосфатаза активируется ионами кальция или инсулином, отщепляет фосфат, активирует пируватдегидрогеназу.
Таким образом, работа пируватдегидрогеназы подавляется при избытке в митохондрии (в клетке) ацетил-SКоА и НАДН, что позволяет снизить окисление пирувата и, следовательно, глюкозы в случае когда энергии достаточно.
Если АТФ мало или имеется влияние инсулина, то образуется ацетил-SКоА. Последний в зависимости от условий будет направляться либо в ЦТК, либо на синтез ХС и ЖК.
6. Цикл трикарбоновых кислот. Общая схема, энергетическое значение
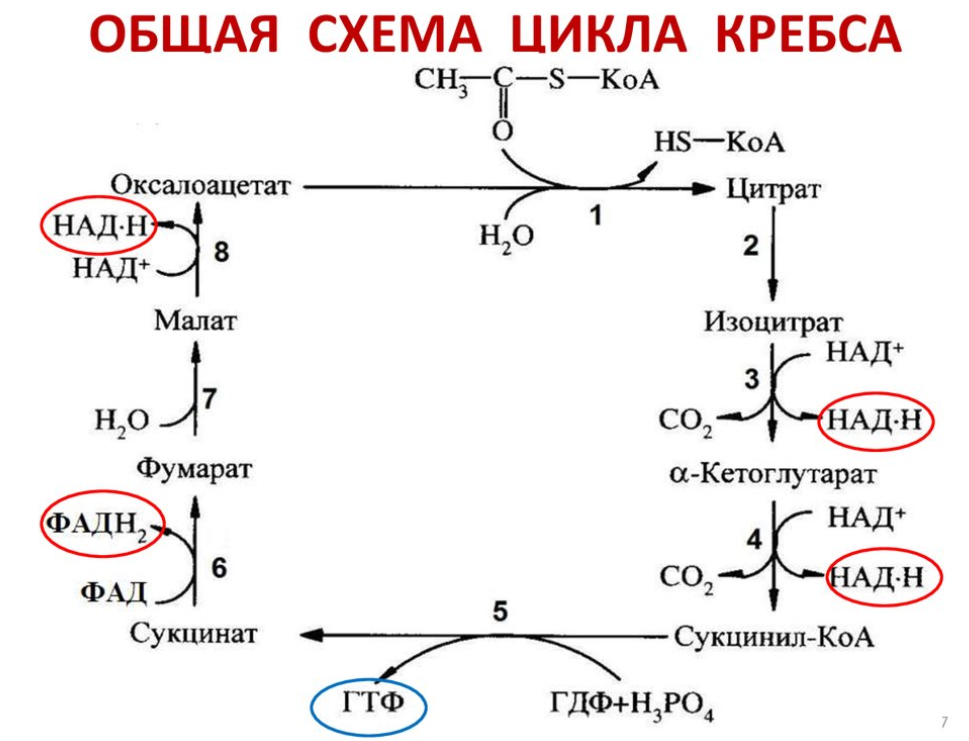
Энергетическое значение:
1) Образуется ГТФ = 1АТФ
2) 3НАДН и 1ФАДН2 - источники водорода для дыхательной цепи, т.е. окислительного фосфорилирования
7. Последовательность реакций цикла трикарбоновых кислот. Реакции дегидрирования. Субстратное фосфорилирование.
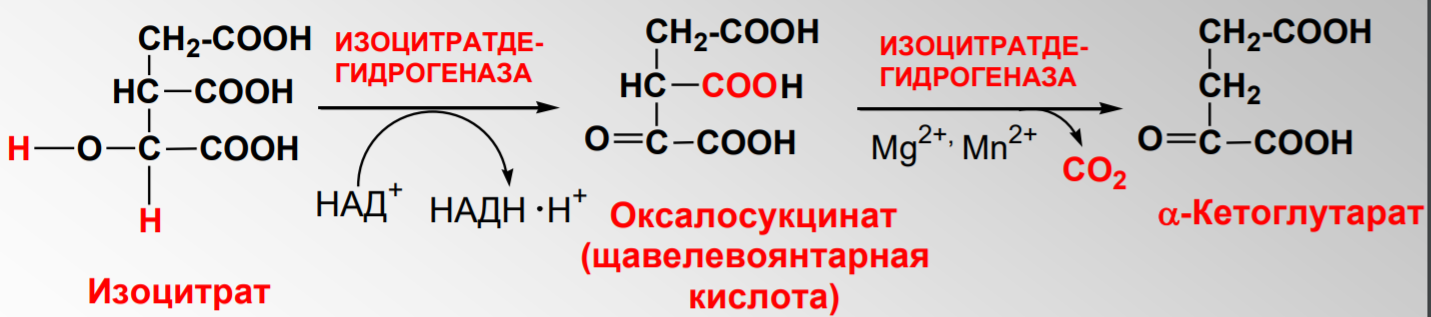
Дегидрирование
Дегидрирование и декарбоксилирование изоцитрата НАД+ зависимой изоцитратдегидрогеназой с образованием a-кетоглутарата:
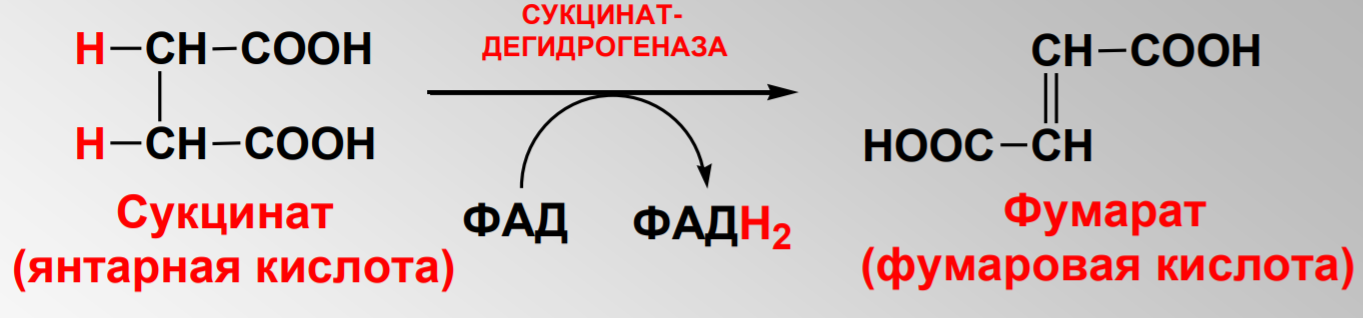
Дегидрирование сукцината до фумарата:
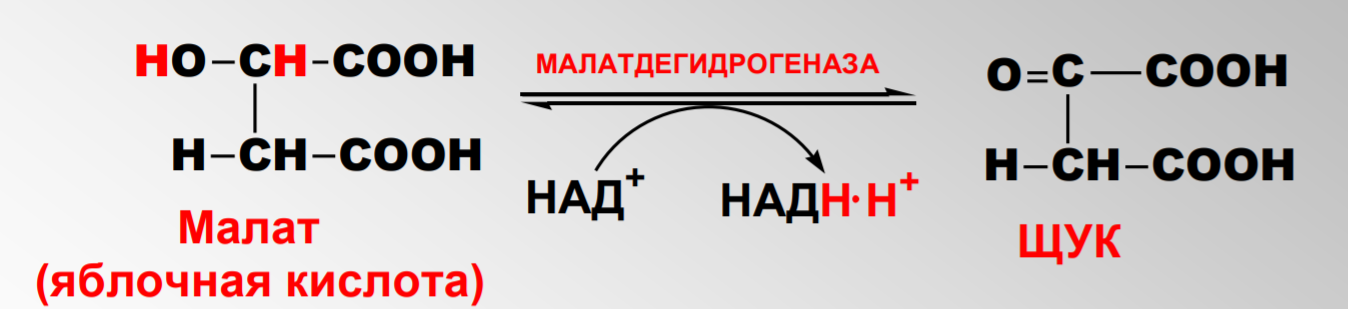
Дегидрирование малата
Окислительное фосфорилирование
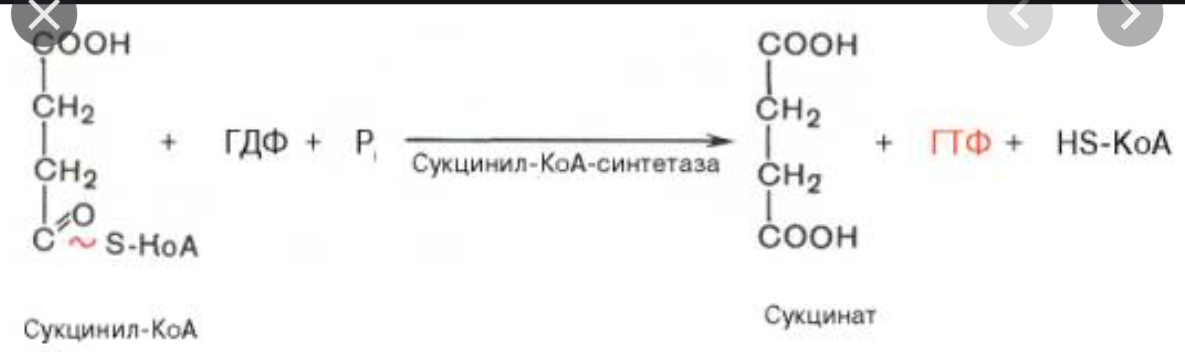
0
0
8. Регуляция цикла трикарбоновых кислот. Анаболические функции цикла трикарбоновых кислот. Анаплеротические реакции
Регуляция цикла трикарбоновых кислот
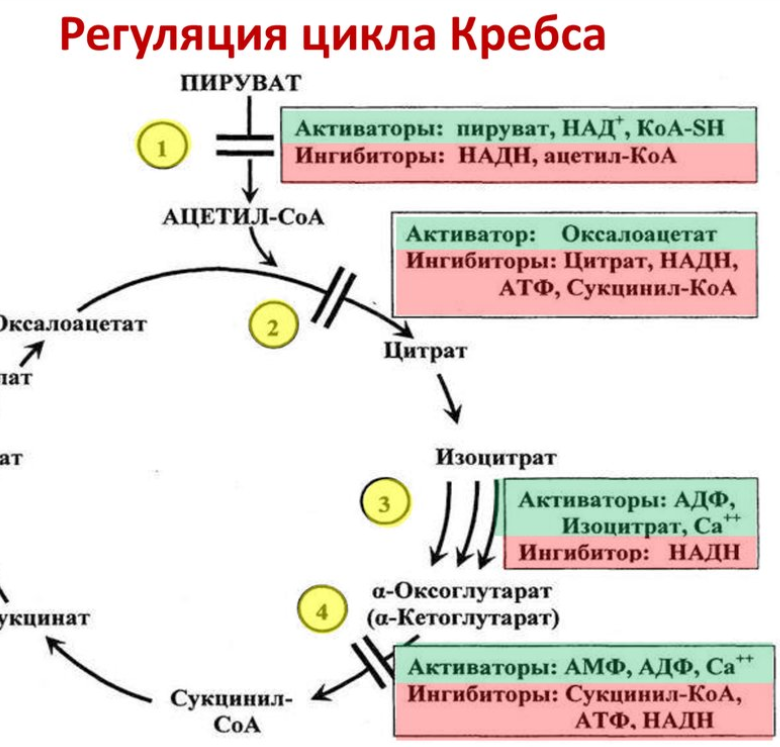
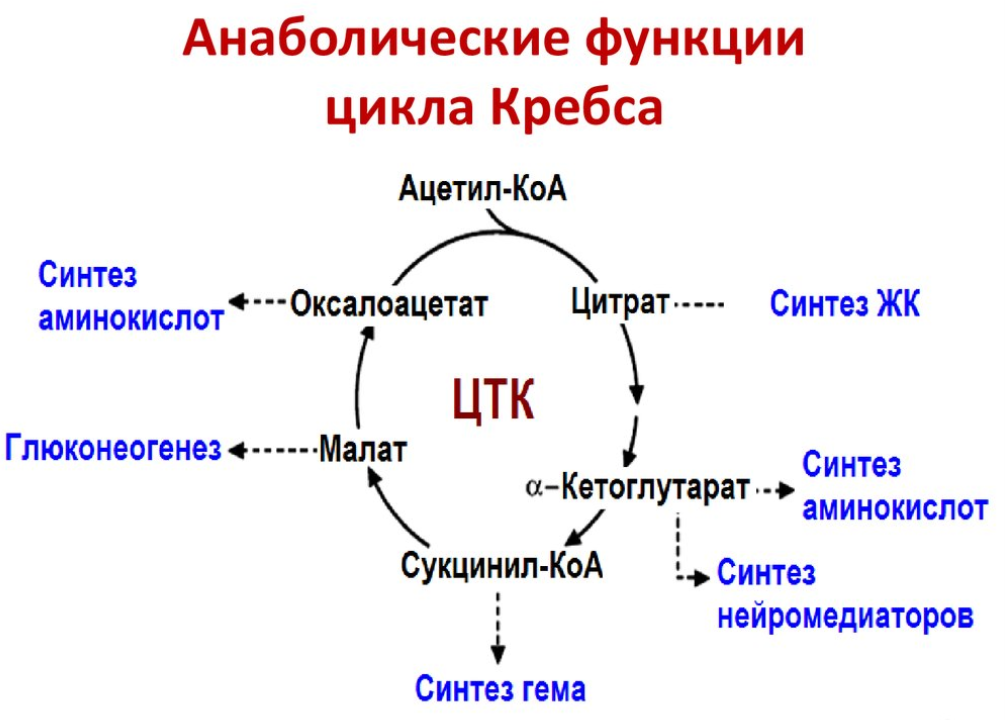
Анаплеротические реакции
Анаплеротические (пополняющие) реакции – специальные ферментативные реакции, обеспечивающие пополнение пула промежуточных продуктов цикла лимонной кислоты.
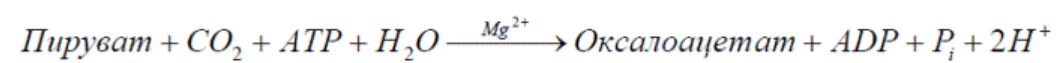
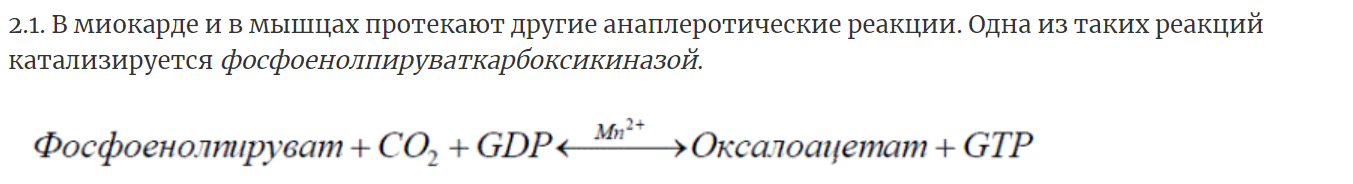

9. Представление о макроэргических субстратах. Классификация макроэргов. Макроэргичность АТФ. АТФ – универсальная энергетическая валюта клетки.
Макроэргические субстраты – биологические молекулы, способные накапливать и передавать энергию. Содержат макроэргические связи, при разрыве которых высвобождается энергия.
Классификация
Их классифицируют по типу связи:
1) фосфоангидридная связь – нуклеозидтри- и нуклеозиддифосфаты (АТФ, ГТФ, АДФ и др.)
2) тиоэфирная связь – ацетил-производные коэнзима А (ацетил-SKoA, сукцинилSKoA)
3) гуанидинфосфатная – креатинфосфат
4) ацилфосфатная – синтез АТФ в субстратном фосфорилировании (1,3- дифосфоглицериновая кислота)
5) енолфосфатная – фосфоенолпируват
АТФ
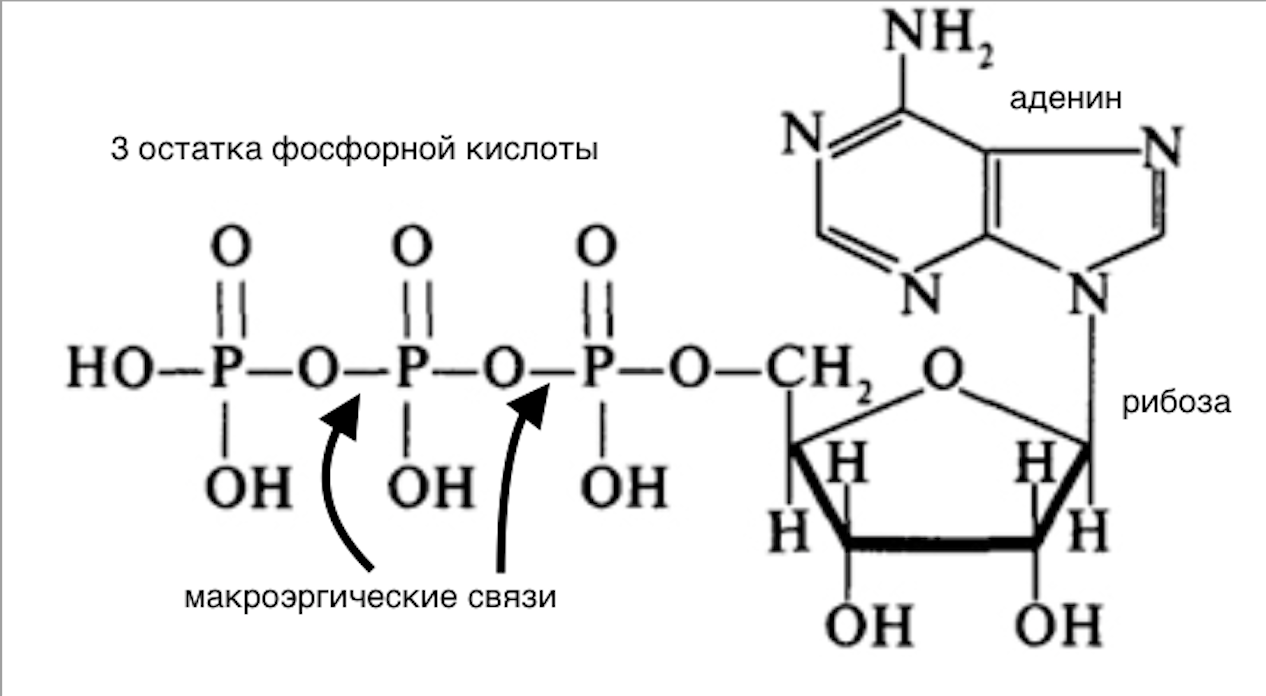
АТФ – макроэргическое соединение, выполняющее функцию аккумулирования биологической энергии и ее переноса для последующего использования при выполнения клеточных функций.
Макроэргичность АТФ объясняется высокой плотностью зарядов, сконцентрированной в «хвосте», которая обеспечивает легкость диссоциации фосфатов при водном гидролизе и высокую величину его свободной энергии.
Молекула АТФ состоит из азотистого основания – аденина, рибозы и трех остатков фосфорной кислоты
10. Пути синтеза АТФ: субстратное и окислительное фосфорилирование. Примеры реакций субстратного фосфорилирования.
Окислительное фосфорилирование – многоэтапный процесс во внутренней мембране митохондрий, происходит окисление восстановленных НАДН и ФАДН2 и сопровождающийся синтезом АТФ.
Субстратное фосфорилирование – фосфорилирование АДФ до АТФ. Передача макроэргического фосфата или энергии макроэргической связи на АДФ.
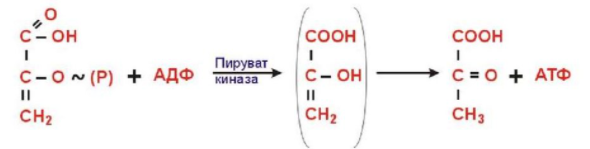
Фосфоенолпируват + АДФ -> енольная форма пирувата -> пируват
11. Окислительное фосфорилирование АДФ. Механизм сопряжения окисления и фосфорилирования. Хемиосмотическая теория Митчелла.
Окислительное фосфорилирование – это многоэтапный процесс, происходящий во внутренней мембране митохондрий и заключающийся в окислении восстановленных эквивалентов (НАДН и ФАДН2) ферментами дыхательной цепи и сопровождающийся синтезом АТФ.
Механизм сопряжения
АТФ образуется путём присоединения к АДФ остатка фосфорной кислоты. Этот процесс называется фосфорилированием. Таким образом, два процесса: процесс биологического окисления (передача протонов и электронов по дыхательной цепи) и процесс фосфорилирования (образование АТФ) являются сопряжёнными, так как энергия, образующаяся при окислении, используется для фосфорилирования. Поэтомуобразование АТФ за счёт энергии, выделяющейся при прохождении электронов по дыхательной цепи, называется окислительным фосфорилированием.
Хемиосмотическая теория Митчелла.
Впервые механизм окислительного фосфорилирования был предложен Питером Митчеллом. Согласно этой гипотезе перенос электронов, происходящий во внутренней митохондриальной мембране, вызывает выкачивание ионов Н+ из матрикса митохондрий в межмембранное пространство. Это создает градиент концентрации ионов Н+ между цитозолем и замкнутым внутримитохондриальным пространством. Ионы водорода в норме способны возвращаться в матрикс митохондрий только одним способом – через специальный фермент, образующий АТФ – АТФ-синтазу.
Центральный постулат - электронпереносящие цепи митохондрий, хлоропластов и бактерий сопряжены с системой синтеза АТФ через разность электрохимических потенциалов протонов на сопрягающих мембранах.
Т.о. АТФ синтезируется за счет кинетической энергии протона, проходящего через АТФ-синтетазу.
12. Состав, структура и номенклатура дыхательных комплексов и других компонентов цепи переноса электронов, их локализация и функции во внутренней мембране митохондрий. Строение АТФ-синтазы. Механизм функционирования.
Всего цепь переноса электронов включает в себя разнообразные белки, которые организованы в 4 больших мембраносвязанных мульферментных комплекса. Также существует еще один комплекс, участвующий не в переносе электронов, а синтезирующий АТФ.
Состав, структура и номенклатура дыхательных комплексов
1 комплекс. НАДН-КоQ-оксидоредуктаза (НАДН-дегидрогеназа)
Структура:
ФМН, 42 белковых молекулы, из них не менее 6 железосерных белков
Функция
- Принимает электроны от НАДН и передает их на коэнзим Q (убихинон).
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
2 комплекс. ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует, его выделение условно. К нему относятся ФАД-зависимые ферменты, расположенные на внутренней мембране:
- ацил-SКоА-дегидрогеназа (β-окисление жирных кислот)
- сукцинатдегидрогеназа (цикл трикарбоновых кислот)
- фосфат-дегидрогеназа (челночный механизм переноса атомов водорода)
Функция
- Восстановление ФАД в окислительно-восстановительных реакциях.
- Обеспечение передачи электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q
3 комплекс. КоQ-цитохром c-оксидоредуктаза (цитохром с редуктаза)
Строение: молекулы цитохрома b и цитохрома c1, железо-серные белки. 2 мономера с 11 полипептидными цепями.
Функция
- Принимает электроны от коэнзима Q и передает их на цитохром с.
- Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
4 комплекс. Цитохром с-кислород-оксидоредуктаза (цитохромоксидаза)
Строение: 13 субъединиц, включает цитохромы а и а3
Функция
- Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
5 комплекс. АТФ-синтаза
Строение: множество белковых цепей, разделенных на 2 группы:
- cубъединица F0, каналообразующая, по ней выкачанные наружу протоны водорода устремляются в матрикс
- cубъединица F1, каталитическая, используя энергию протонов, синтезирует АТФ
Механизм работы АТФ-синтазы получил название вращательный катализ:
- синтез АТФ происходит за счет энергии, высвобождающейся при прохождении протонов через каналы комплекса Fo
- протоны из межмембранного пространства достигают своих центров связывания на с-субъединицах через входной полуканал и прикрепляются к аспартату, нейтрализуя его отрицательный заряд -> лишенная заряда с-субъединица меняет свою конформацию и принуждает Fo-комплекс вращаться вокруг своей оси, доставляя протоны к другому полуканалу, который направлен уже в матрикс митохондрий -> поскольку концентрация ионов H+ в матриксе низка, то они легко отрываются от аспартата и уходят внутрь, а оставшийся, уже заряженный, аспартат не позволяет комплексу вращаться в обратном направлении.
-при связывании ионов H+ происходит поворот не только комплекса c-субъединиц Fo-компонента, но и жестко связанной с ним γ-субъединицы F1-компонента -> стержень γ–субъединицы проворачивается и при каждом повороте на 120° она поочередно вступает в контакт с каталитическими β-субъединицами, что меняет их функционирование -> при каждом обороте γ-субъединицы на 360º синтезируются три молекулы АТФ.
13. Регуляция окислительного фосфорилирования. Дыхательный контроль. Механизмы разобщения окисления и фосфорилирования. Физиологическое значение разобщения. UCP-белки. Молекулярно-биологические аспекты функции и дисфункции митохондрий
Регуляция окислительного фосфорилирования. Дыхательный контроль.

Механизмы разобщения окисления и фосфорилирования
Для того, чтобы окислительное фосфорилирование происходило нужна абсолютная непроницаемость внутренней мембраны митохондрий для протонов, иначе нельзя будет создать протонный градиент.
Значит вещества, нарушающие целостность митохондриальной мембраны, препятствуют возникновению протонного градиента и разобщают процессы окисления и фосфорилирования.
При этом возрастает доля свободного окисления, и вся энергия, освобождающаяся в процессе транспорта электронов, выделяется в виде тепла.
Поэтому все разобщители обладают пирогенным действием, т.е. повышают температуру тела.
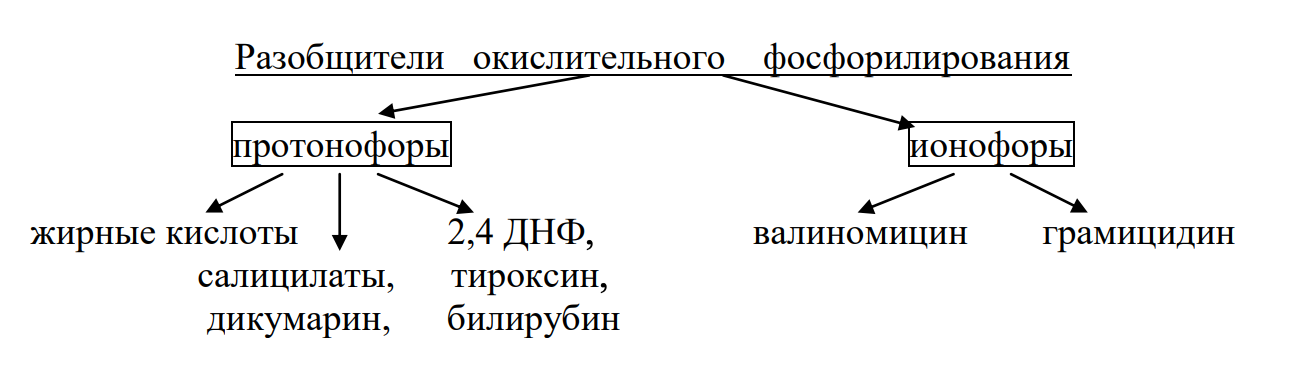
Разобщители:
1) Протонофоры - переносят Н+ - ионы через мембраны в обход протонных каналов. (должны уметь проходить через био мембраны - липофильные)
Так, жирные кислоты, которые в форме аниона R-СОО- связывают протоны на внешней стороне мембраны, транспортируют их в виде недиссоциированной формы (R-СООН) внутрь митохондрии, где диссоциируют, освобождая ионы Н+ и таким образом ликвидируют протонный градиент.
2) Ионофоры - вещества, повышающие проницаемость мембраны для одного из катионов (Na+ или K + ) или для Н+ и катиона.
Антибиотик валиномицин, связывает ионы калия на наружной стороне митохондриальной мембраны, переносит их внутрь митохондрий, чем полностью компенсируется разность зарядов на различных сторонах мембраны.
Биологическое значение
Разобщение происходит при лихорадочных состояниях (некоторые токсины бактерий), при понижении температуры тела (холодовое разобщение), авитаминоз Е
В ряде случаев разобщение может быть биологически целесообразным, так как является способом генерирования тепла для поддержания температуры тела у зимнеспящих животных, у новорожденных и у млекопитащих, адаптированных к холоду
UCP - белки
- термогенин
- обнаружен в митохондриях адипоцитов бурой жировой ткани
- основной механизм теплопродукции и новорожденных и животных в спячке
- кодируется геномом
- увеличивают проницаемость внутренней митохондриальной мембраны, позволяя протонам, перенесенным в межмембранное пространство, возвращаться в митохондриальный матрикс.
14. Активные формы кислорода и азота. Неферментативные и ферментативные пути образования. Перекисное окисление липидов: механизм процесса, продукты. Первичные и вторичные механизмы повреждающего действия окислительного стресса. Перекисная теория гибели клетки
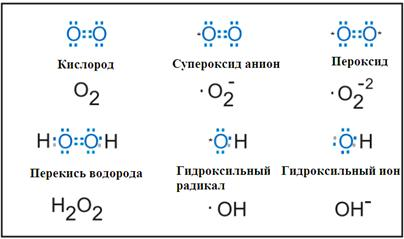
Активные формы азота
Пероксинитрит анион - ONOO-
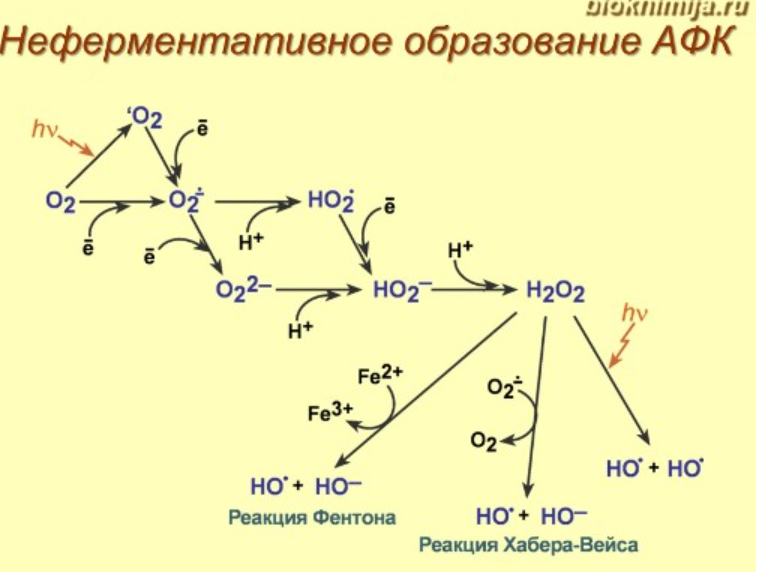
См. тетрадь
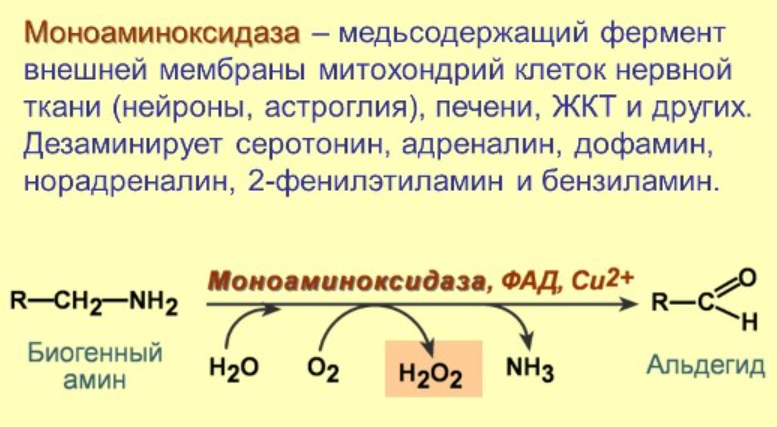
Ферментативные пути образования:
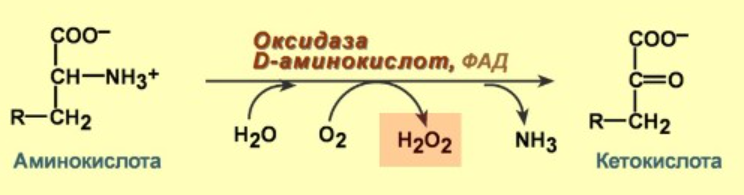
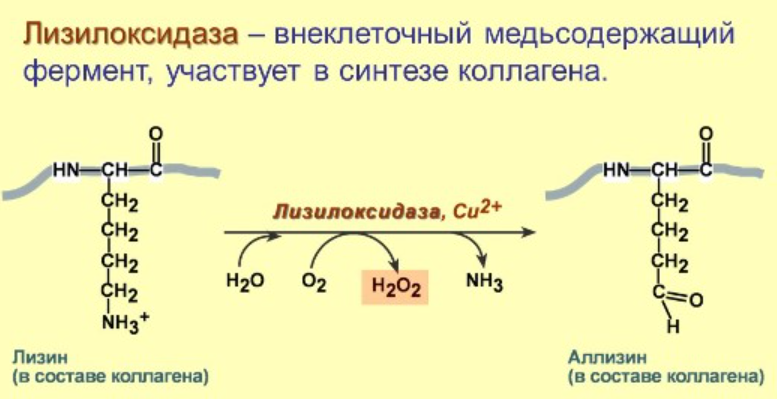
Перекисное окисление липидов: механизм процесса, продукты
При окислительном стрессе лидирует окисление жирнокислотных остатков в фосфолипидах мембран. Это снижает их гидрофобность и нарушает устойчивость мембран, изменяет работу мембраносвязанных ферментов, повышает проницаемость мембран для ионов.
Перекисное окисление липидов включает в себя несколько стадий:
- Инициация - атакуется метиленовая группа, расположенная между двойными связями, и выбивается атом водорода, восстанавливающий гидроксил-радикал до воды. Далее в жирной кислоте происходит перестановка двойной связи, смещение радикальной группы и взаимодействие ее с кислородом. В результате образуется липопероксильный радикал
- Развитие - дальнейшее взаимодействие полученного липопероксильного радикала с соседними жирными кислотами приводит к его нейтрализации и появлению новых липоперекисных радикалов, т.е. к развитию линейной цепной реакции с появлением новых окисленных жирных кислот.
- Разветвление - кроме линейного развития, может происходить ветвление реакции за счет получения гидроперекисью электронов от каких-либо металлов или при воздействии излучения
- Обрыв цепи - обрыв цепной реакции происходит при взаимодействии радикалов друг с другом или в реакции с различными антиоксидантами, например, витамином Е, который отдаёт электроны, превращаясь при этом в довольно стабильную окисленную форм
Продукты:
Первичные - гидроперекиси жирных кислот
Вторичные (от распада гидроперекиси) - спирты, кетоны, альдегиды. Запомнить - малоновый диальдегид (наиболее реакционноспособный)
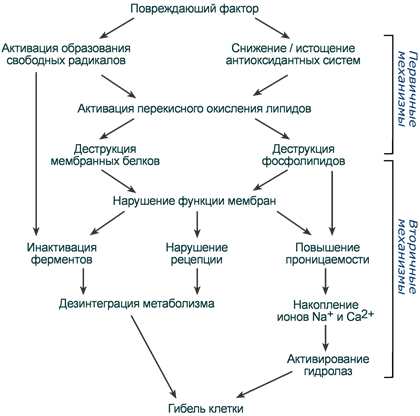
Первичные и вторичные механизмы повреждающего действия окислительного стресса
Окислительный стресс - избыточная продукция эндогенных АФК или из-за большой концентрации экзогенных АФК
Первичные м-мы:
Процессы окисления непосредственно в местах своего образования – митохондриях, клеточной мембране, мембранах ЭПР. Под влиянием свободных радикалов происходит окисление молекул – ядерной и митохондриальной ДНК, РНК, белков, фосфолипидов и гликозаминогликанов
Вторичные м-мы:
Деструкция мембран, внутриклеточных структур, органелл
Свободнорадикальная теория старения утверждает, что старение происходит из-за накопления повреждений в клетках, нанесённых свободными радикалами с течением времени
Свободнорадикальная митохондриальная теория старения, представляющая собой модификацию свободнорадикальной теории старения, утверждает, что старение у людей и животных обусловлено накоплением повреждений в митохондриях и митохондриальной ДНК[
15. Антиоксидантная система организма. Антиоксиданты неферментативной природы. Ферментные системы антиоксидантной защиты.
Живые клетки имеют систему защиты от повышенной продукции свободных радикалов - антиоксидантную систему, механизм действия которой направлен на блокирование действия свободных радикалов.
Антиоксидантная система образована антиоксидантными ферментами низкомолекулярными антиоксидантами. Антиоксиданты «ловят» радикалы, отдавая им свои электроны, таким образом, обезвреживая их. При этом структура антиоксидантов остается стабильной.
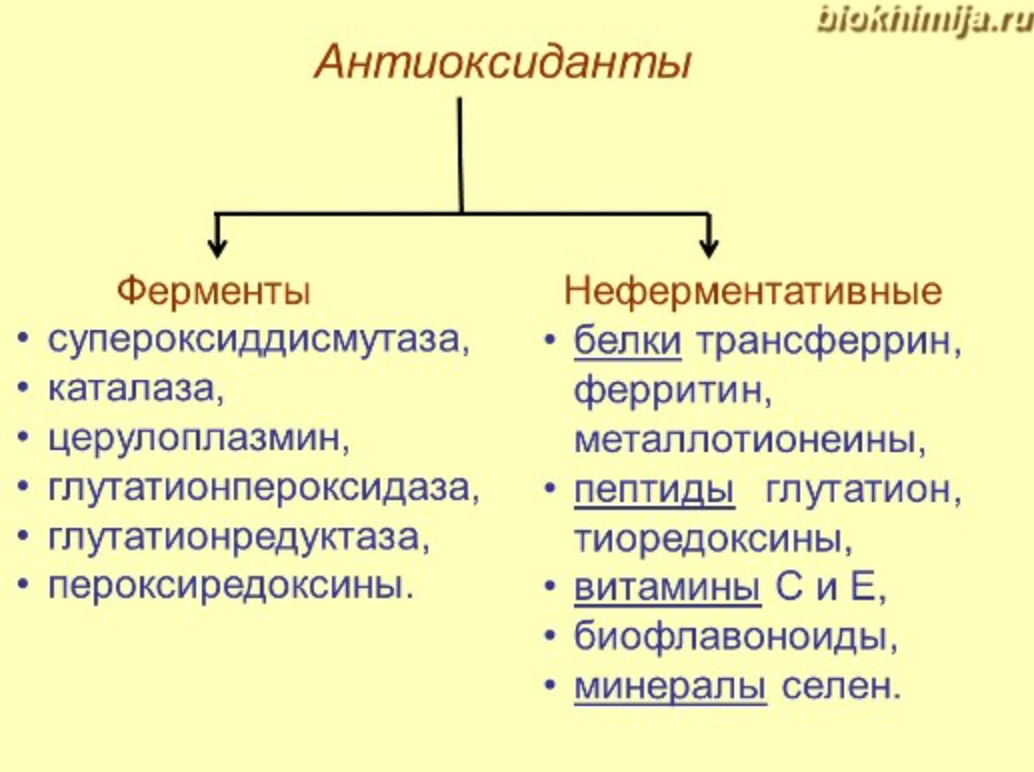
Антиоксиданты неферментативной природы
1) Ферритин и трансферрин - способны связывать ионы железа и предотвращать реакцию Фентона
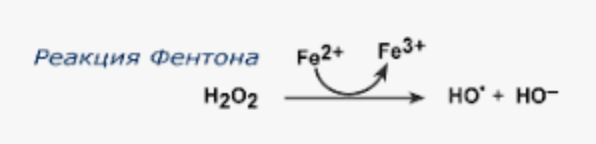
2) Металлопротеины - богаты цистеином, способны связывать ионы металлов переменной валентности - железо, цинк, медь
3) Токоферол
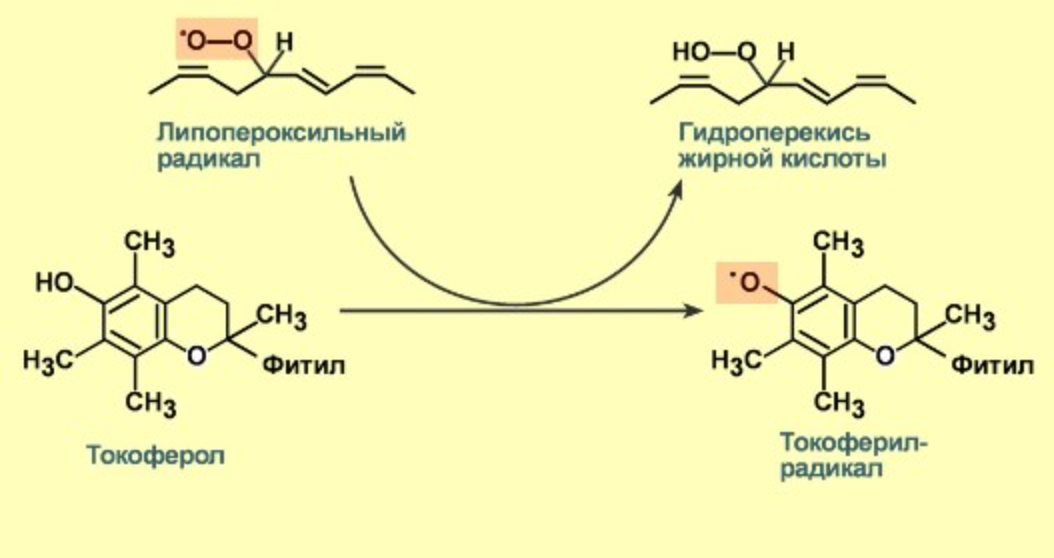

Так в принципе у многих витаминов
Ферментные системы антиоксидантов
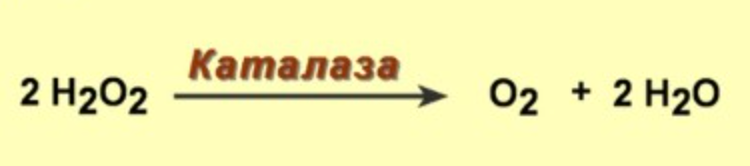
1) Супероксиддисмутаза - фермент цитозоля и митохондрий, из радикалов образуется перекись

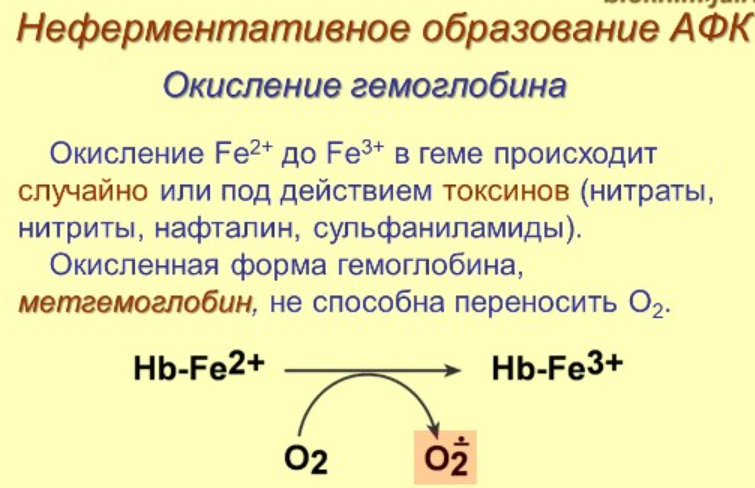
2) Дальше действует каталаза, содержит гем, есть во всех пероксисомах и в цитозоле эритроцитов - защищает гемоглобин от окисления
3) Церулоплазмин - альфа глобулин, содержит Cu, ускоряет связывание железа с трансферрином
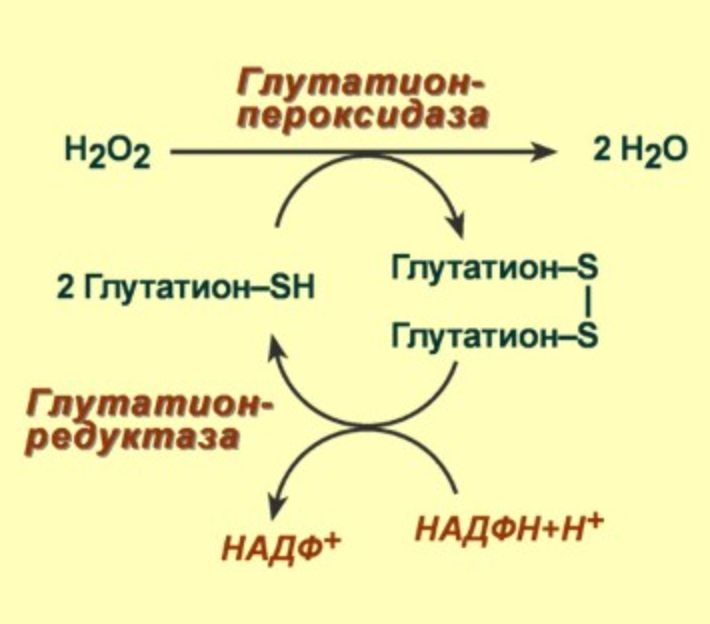
4) Глутатионпероксидаза, содержит гем, в 1000 раз большее сродство к перекиси, чем у каталазы (есть селеноцистеин в активном центре - цистеин с селеном вместо серы)
16. Физиологическое значение свободно-радикального окисления. Свободнорадикальное окисление, фагоцитоз и воспаление. Свободнорадикальный механизм антимикробной защиты грудного молока. Свободнорадикальное окисление при патологии сердечно-сосудистой системы.
Физиологическая роль свободных радикалов достаточно велика. Большая часть свободных радикалов генерируются фагоцитами, Т-лимфоцитами при воспалительных реакциях и выполняют защитную роль, лизируя патогенные микроорганизмы, мутировавшие (раковые) клетки.
Важнейшую физиологическую роль выполняет генерируемый в эндотелии сосудов оксид азота NO (эндотелий-зависимый релаксирующий фактор), обеспечивающий расслабление гладкой мускулатуры сосудистой стенки и регулирующий уровень артериального давления, коронарный и органный кровоток, а также предотвращающий агрегацию тромбоцитов
Свободнорадикальное окисление, фагоцитоз и воспаление
Фагоцитоз – это активное поглощение и разрушение клетками твердых частиц, чуждых для организма (микроорганизмы, вирусы, иммунные комплексы антиген-антитело, простейшие, гельминты, отмершие клетки, твердые неорганические частицы)
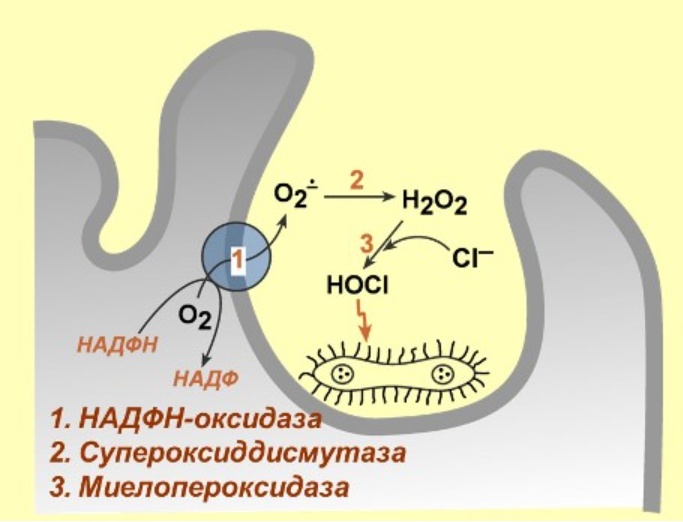
Свободнорадикальный механизм антимикробной защиты грудного молока


Свободнорадикальное окисление при патологии сердечно-сосудистой системы
При гипоксии (ишемии) миокарда окислительные процессы в митохондриях кардиомиоцитов нарушаются (как бы не доходят до конца), в результате чего накапливаются промежуточные метаболиты цикла Кребса, крайне легко подверженные восстановлению с образованием свободных радикалов и перекисных соединений, угнетающих систему антиоксидантной защиты. В конечном итоге создается парадоксальная ситуация — уменьшение кислорода в клетке приводит к увеличению кислородных радикалов. Развивающаяся после каждого эпизода транзиторной ишемии реперфузия миокарда также сопровождается значительной активацией (в сотни раз) свободнорадикальных процессов и выбросом липопероксидов в кровоток.
Велика роль свободнорадикального окисления (СРО) в патогенезе нестабильной стенокардии и инфаркта миокарда. Локальная активация СРО в зоне ишемии и накопление продуктов деградации свободных радикалов стимулируют свертываемость крови, увеличивают ее вязкость, усиливают агрегацию и адгезию форменных элементов крови. Высокая концентрация пероксидов ускоряет дегенерацию NO с образованием пероксинитрита — крайне цитотоксичного соединения: О2– +NOо ->ОNOO–. Ускоренный распад эндотелиального NO стимулирует ангиоспазм, а окисление экзогенного NO, образующегося в результате метаболизма потребляемых больным нитропрепаратов, уменьшает их терапевтическую эффективность. Кроме того, свободные радикалы модифицируют эндотелиальные NO-рецепторы, уменьшая их чувствительность, а также оказывают непосредственное повреждающее действие на кардиомиоциты. Указанные процессы усугубляют ишемию, оказывают аритмогенный эффект, способствуют распространению зоны некроза и повреждения.