Биохимия углеводы
1. Биологическая роль углеводов. Суточная потребность в углеводах у взрослых и детей. Углеводы пищи животного и растительного происхождения, их значение.
Биологическая роль

Суточная потребность
Взрослые - 450-550 г/сут
Дети: для детей до года 13 г/кг массы тела, для детей старше года от 170 до 420 г/сутки.
В пище человека в основном содержатся полисахариды — крахмал, целлюлоза (растений), в меньшем количестве - гликоген (животных).
Источником сахарозы служат растения, особенно сахарная свёкла, сахарный тростник.
Лактоза поступает с молоком млекопитающих (в коровьем молоке до 5% лактозы, в женском молоке — до 8%).
Фрукты, мёд, соки содержат небольшое количество глюкозы и фруктозы.
Мальтоза есть в солоде, пиве.
Углеводы пищи являются для организма человека в основном источником моносахаридов, преимущественно глюкозы.
Некоторые полисахариды: целлюлоза, пектиновые вещества, декстраны, у человека практически не перевариваются, в ЖКТ они выполняют функцию сорбента (выводят холестерин, желчные кислоты, токсины), необходимы для стимуляции перистальтики кишечника и формирования нормальной микрофлоры.
Функции хз надо ли
1. Энергетическая функция. при окислении 1 грамма углеводов выделяются 4,1 ккал энергии и 0,4 г воды.
2. Пластическая функция. Углеводы входят в состав сложных молекул (например, пентозы (рибоза и дезоксирибоза) участвуют в построении АТФ, ДНК и РНК).
3. Запасающая функция. Углеводы выступают в качестве запасных питательных веществ: гликоген у животных, крахмал и инулин — у растений.
4. Осмотическая функция. Углеводы участвуют в регуляции осмотического давления в организме. Так, в крови содержится 100—110 мг/л глюкозы, от концентрации которой зависит осмотическое давление крови.
5. Рецепторная функция. Олигосахариды входят в состав воспринимающей части многих клеточных рецепторов или молекул-лигандов.
3. Переваривание углеводов. Характеристика и действие ферментов участвующих в полостном и пристеночном пищеварении: α-амилаза ротовой полости, ферменты панкреатического сока, ферментные комплексы тонкого кишечника, отвечающие за гидролиз дисахаридов.
Переваривание происходит за счет ферментов пищеварительных соков:
Ротовая полость: В слюне содержится альфа-амилаза, она беспорядочно разрушает альфа-1,4 гликозидные связи, не трогая другие, получаются декстрины (некоторые из них могут перевариться до мальтозы и изомальтозы)
Дисахариды не гидролизуются
Желудок: pH желудка инактивирует амилазу, но если пищевой комок хорошо сформирован, то в его глубине может продолжаться действие фермента
Кишечник: При полостном пищеварении работает альфа-амилаза панкреатического сока, разрывающая в ранее образованных (слюнной амилазой) декстринах, в крахмале и гликогене внутренние α1,4-связи, переваривая до мальтозы и декстринов
При пристеночном пищеварении работают ферментные комплексы:
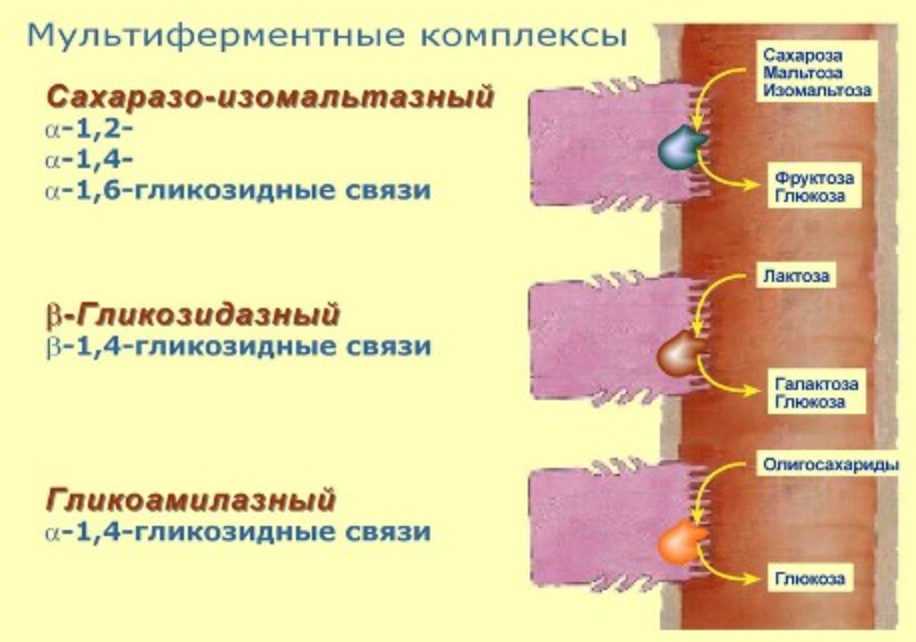
- сахаразо-изомальтазный комплекс (рабочее название сахараза) – в тощей кишке гидролизует альфа-1,2-, альфа1,4-, альфа-1,6-гликозидные связи, расщепляет сахарозу, мальтозу, мальтотриозу, изомальтозу
- бета-гликозидазный комплекс (рабочее название лактаза) – гидролизует бета1,4-гликозидные связи между галактозой и глюкозой (лактозу).
У детей активность лактазы очень высока уже до рождения и сохраняется на высоком уровне до 5-7 лет, после чего снижается, - гликоамилазный комплекс – находится в нижних отделах тонкого кишечника и расщепляет альфа-1,4-гликозидные связи в олигосахаридах с восстанавливающего конца.
5. Транспорт моносахаридов через клеточные мембраны: облегченная диффузия и активный транспорт. Транспортёры глюкозы: виды, особенности структуры, функции. Инсулин-зависимые транспортеры.
Всасывание в кишечнике:
Вторично-активный транспорт, но энергия тратится не на сахар, а на создание химического градиента натрия.
Na+ ,К+ -АТФаза постоянно выносит из клетки натрий в обмен на калий -> получается в просвете много натрия -> но есть специальный мембранный белок, у которого есть два центра связывания -> один для натрия, второй для моносахарида -> (причем моносахарид связывается только после натрия) -> белок ползет внутрь клетки -> как только контактирует с цитоплазмой натрий отвязывается по градиенту концентрации, вместе с ним улетает моносахарид -> натрий выкачивается АТФ-азой опять
Выход в кровь
Выход глюкозы из клетки в межклеточное пространство и далее кровь происходит благодаря облегченной диффузии.
Из крови в клетки:
Облегченная диффузия по градиенту концентрации через белки-переносчики
В печени и эпителии кишечника присутствует ГлюТ-2, всегда присутствующий на мембране и обладающий способностью пропускать глюкозу в обе стороны – как внутрь клетки, так и наружу.
В мышцах и жировой ткани находится ГлюТ-4, только эти транспортеры являются чувствительными к влиянию инсулина – при действии инсулина на клетку они поднимаются к поверхности мембраны и переносят глюкозу внутрь. Именно поэтому данные ткани получили название инсулинзависимых.
Некоторые ткани совершенно нечувствительны к действию инсулина, их называют инсулиннезависимыми. К ним относятся нервная ткань, стекловидное тело, хрусталик, сетчатка, клубочковые клетки почек, эндотелиоциты, семенники и эритроциты.
Большинство клеток занимает промежуточное положение, т.е. на их мембранах находятся разные типы транспортеров.
Глют 3 - нейроны и плацента - высокое сродство к глюкозе, работает даже при оч низких концентрациях
Глют 1- в зародышевых тканях - экспрессируется при низких концентрациях
7. Особенности ферментов обмена углеводов в печени: участие глюкокиназы и глюкозо-6-фосфатазы в поддержании постоянной концентрации глюкозы в крови.
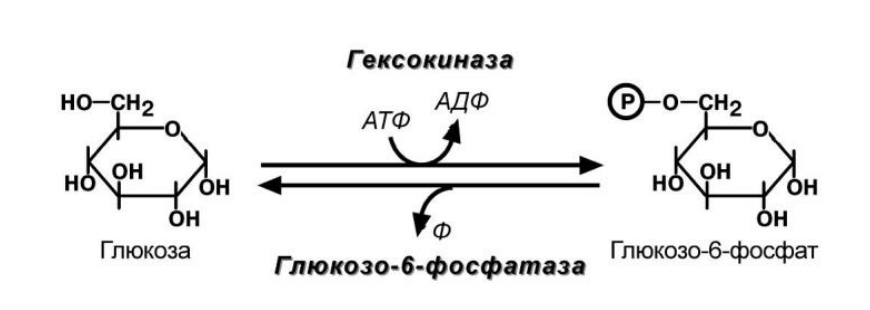
Глюкокиназа
- Глюкокиназа имеет низкое сродство к глюкозе, работает только при высоких концентрациях -> печень не будет использовать глюкозу, если её концентрация в крови в нормальном диапазоне
- Изофермент гексокиназы - гексокиназа 4
- Продукт реакции (глюкозо-6-фосфат) не ингибирует фермент, в то время как в других тканях гексокиназа чувствительна к такому влиянию. Это позволяет гепатоциту в единицу времени захватывать глюкозы больше, чем он может сразу же утилизовать
- Чувствительна к действию инсулина - активируется им
Благодаря таким отличиям гепатоцит может эффективно захватывать глюкозу после еды, накапливать глюкозо-6-фосфат и, "не торопясь", метаболизировать его в любом направлении – синтез гликогена, ПФП, окисление до ацетил-SКоА, CO2 и H2O, и синтез липидов.
Глюкозо-6-фосфатаза
- дефосфорилирование глюкозы осуществляется глюкозо-6-фосфатазой
- есть только в печени и почках - поэтому только они могут продуцировать свободную глюкозу
- в гепатоцитах поддерживает постоянный уровень глюкозы в крови при голодании и мышечных нагрузках
- образуемая свободная глюкоза уходит через плазматическую мембрану в кровь
- фермент последней реакции глюконеогенеза
Только печень работает на обеспечение глюкозой (мобилизацию гликогена) другими органами, остальные используют гликоген только для своих нужд
9. Распад гликогена до глюкозо-6-фосфата. Биологическое значение, реакции, ферменты. Тканевая и клеточная локализация.
Распад (мобилизация) гликогена - гликогенолиз - активируется при недостатке свободной глюкозы в клетках, а значит и в крови
Опять же:
Только печень работает на обеспечение глюкозой (мобилизацию гликогена) другими органами, остальные используют гликоген только для своих нужд
В гликогенолизе непосредственно участвуют три фермента:
1. Фосфорилаза гликогена (кофермент пиридоксальфосфат В6) – расщепляет α1,4-гликозидные связи с образованием глюкозо-1-фосфата. Фермент работает до тех пор, пока до точки ветвления α(1-6) не останется 4 остатка глюкозы.
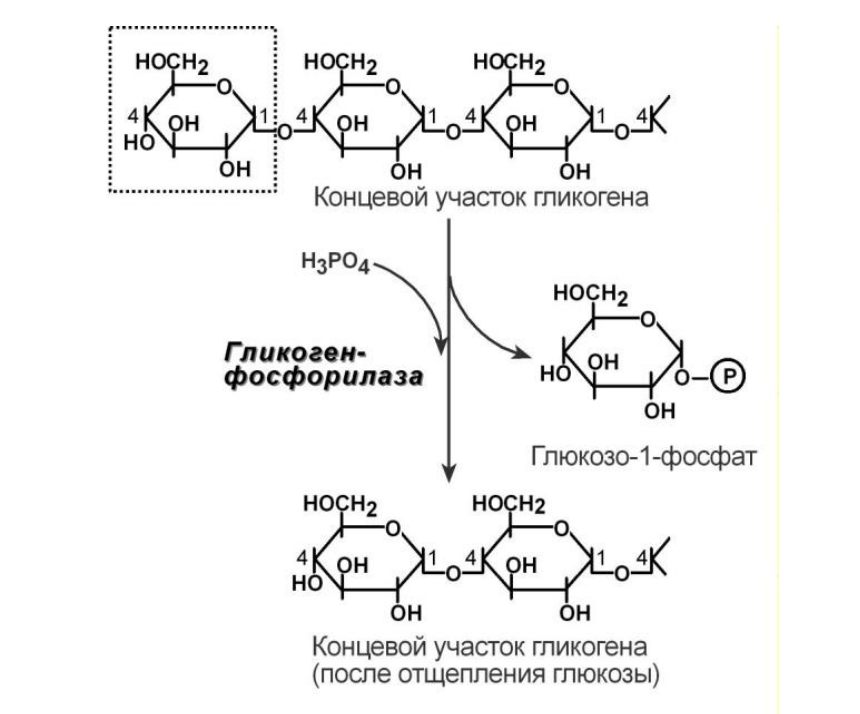
2. α(1-4)-α(1-4)-Глюкантрансфераза – фермент, переносящий фрагмент из трех остатков глюкозы на другую цепь с образованием новой α1,4-гликозидной связи. При этом на прежнем месте остается один остаток глюкозы и "открытая" доступная α1,6-гликозидная связь.

3. Амило-α1,6-глюкозидаза, "деветвящий" фермент – гидролизует α1,6-гликозидную связь с высвобождением свободной (нефосфорилированной) глюкозы. В результате образуется цепь без ветвлений, служащая субстратом для фосфорилазы.
Тканевая и клеточная локализация?????
11.Регуляция ферментов обмена гликогена – гликогенсинтазы и гликоген-фосфорилазы: гормональная – влияние адреналина и глюкагона (аденилатциклазный механизм, роль цАМФ и протеинкиназы А); роль инсулина и участие фосфодиэстеразы в снижении концентрации цАМФ в клетке; аллостерическая регуляция активности гликогенфосфорилазы при участии АМФ; кальций-зависимая активация киназы фосфорилазы гликогена.
Основные ферменты метаболизма гликогена активны либо в фосфорилированной, либо в дефосфорилированной форме.
Присоединение фосфатов к ферментам производят протеинкиназы, источником фосфата является АТФ:
1) Гликогенсинтаза - активна дефосфорилированной
2) Гликоген-фосфорилаза - активна фосфорилированной
Адреналин и глюкагон
Фосфорилируют их вместе - значит вызывают гликогенолиз - тк активируют фосфорилазу
МАЛО ГЛЮКОЗЫ ИЛИ НУЖНО БОЛЬШЕ - АКТИВАЦИЯ ГЛИКОГЕНОЛИЗА, ЧТОБЫ ВЫСВОБОЖДАЛАСЬ ГЛЮКОЗА
Инсулин
Инсулин -> активирует протеинфосфатазу -> дефосфорилирование -> активация синтеза гликогена
ЛОГИЧНО НУЖНО УМЕНЬШИТЬ КОЛИЧЕСТВО ГЛЮКОЗЫ В КРОВИ - АКТИВИРУЕТСЯ ГЛИКОГЕНЕЗ
Одновременно инсулин и глюкокортикоиды (кортизол) увеличивают синтез гликогена, увеличивая количество молекул гликогенсинтазы.
Фосфорилаза гликогена
Скорость распада гликогена лимитируется только скоростью работы фосфорилазы гликогена. Ее активность может изменяться тремя способами:
- ковалентная модификация
- кальций-зависимая активация
- аллостерическая активация с помощью АМФ
Ковалентная модификация
аденилатциклазный механизм, роль цАМФ и протеинкиназы А, роль инсулина и участие фосфодиэстеразы в снижении концентрации цАМФ в клетке
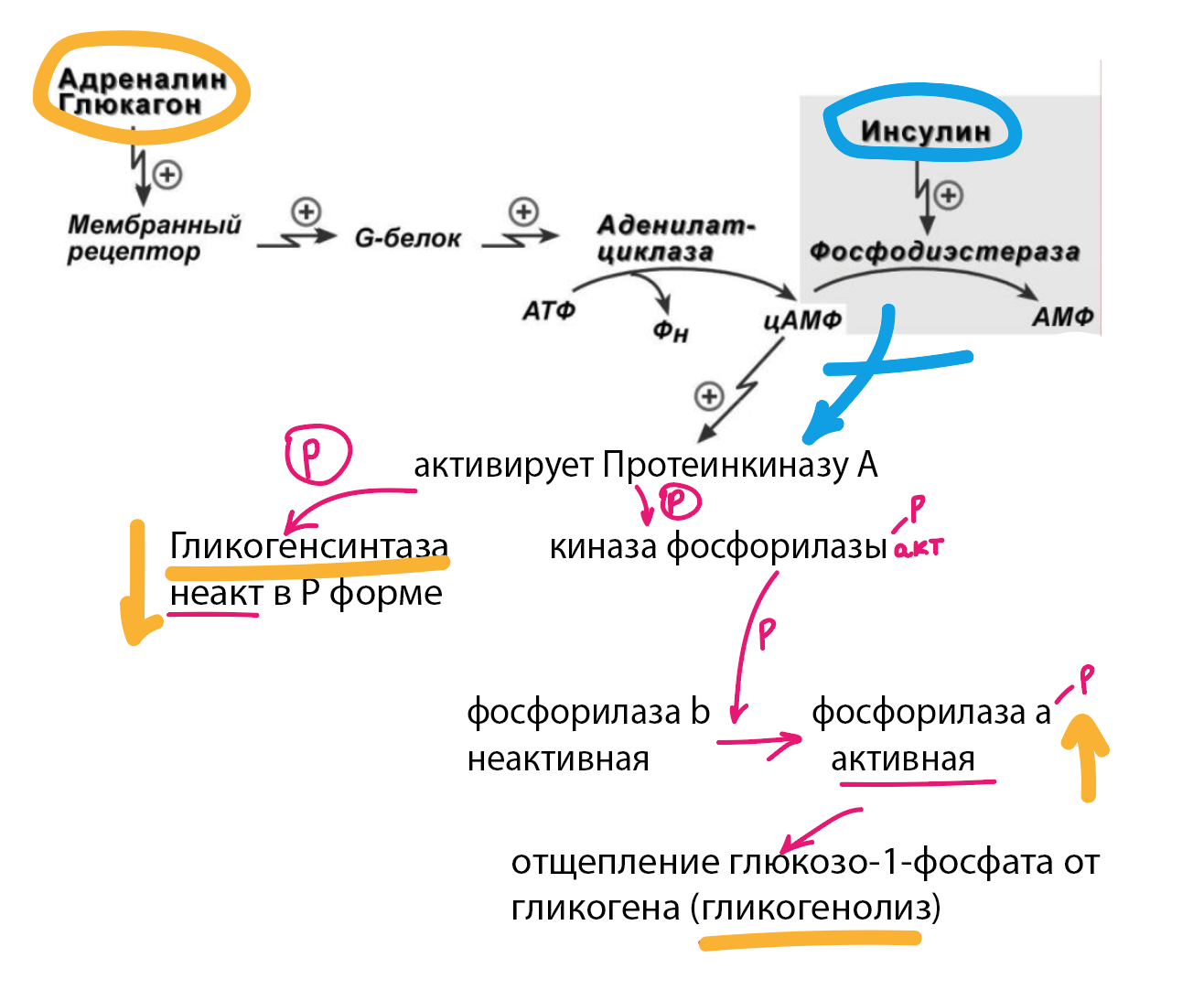
Кальций-зависимая активация
Киназа фосфорилазы активируется не протеинкиназой, а Са и кальмодулином
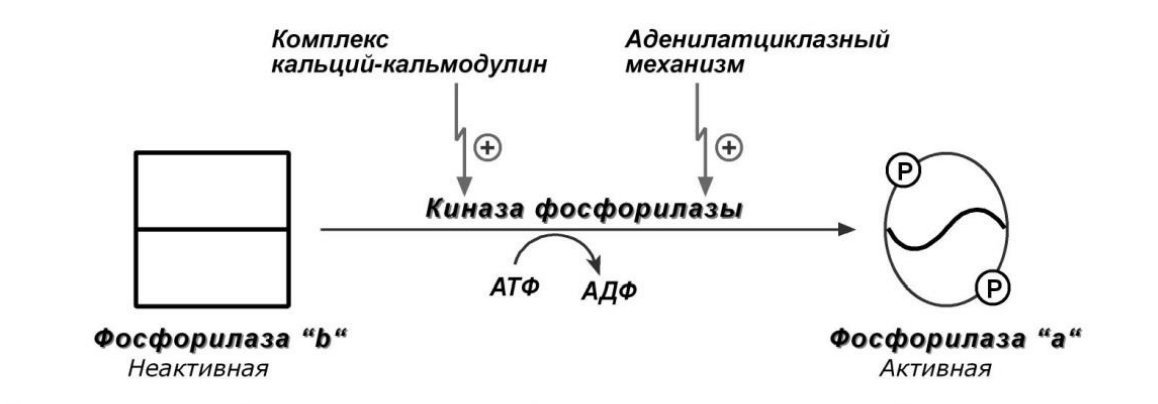
Этот путь работает при работе кальцийфосфолипидного механизма, либо при отсутсвии стимуляции аденилатциклазы.
Это оправдывает себя, например, в мышцах при умеренной нагрузке, когда влияние адреналина через аденилатциклазу еще не произошло или недостаточно, но в цитоплазму под влиянием нервных импульсов поступают ионы Ca2+ .
Аллостерическая регуляция
Также существует активация фосфорилазы гликогена с помощью АМФ – аллостерическая активация благодаря присоединению АМФ к молекуле фосфорилазы "b". Способ работает в любой клетке – при увеличении расхода АТФ и накоплении продуктов его распада АДФ и АМФ.
13.Источники и пути превращения глюкозы в клетке. Роль глюкозо-6-фосфата в метаболизме глюкозы. - см 6 вопрос
15.Характеристика процесса глюконеогенеза: локализация и условия протекания реакций, субстраты, последовательность реакций и ферменты, реакции глюконеогенеза, сопряженные с потреблением ГТФ и АТФ, необратимые реакции глюконеогенеза, значение при голодании и физической работе, расход энергии для синтеза одной молекулы глюкозы. Роль гликолиза и глюконеогенеза в метаболизме плода и новорожденных.
Таким образом, при определенных ситуациях – при низком содержании углеводов в пище, голодании, длительной физической работе – организм должен иметь возможность получить глюкозу. Это достигается процессом глюконеогенеза.
Глюконеогенез - получение глюкозы из других орг. соединений неуглеводного строения.
Последняя реакция может происходить только в печени и почках (только там есть глюкозо-6-фосфатаза), поэтому в полном объеме глюконеогенез происходит только там, остальные реакции могут протекать во всех тканях.
Субстраты: лактат, пируват, глицерол, кетокислоты цикла Кребса и другие кетокислоты, аминокислоты (кроме лейцина и лизина)
17.Анаэробное окисление глюкозы. Судьба продуктов гликолиза в анаэробных условиях. Энергетический эффект окисления глюкозы и гликогена в анаэробных условиях.
19.Этапы аэробного окисления и суммарное уравнение аэробного распада глюкозы. Преимущества аэробного окисления.
21.Характеристика витаминов, участвующих в превращении пирувата (H, В1, В2, В3, В5): источники, коферментные формы, суточная потребность, биохимические функции, характерные признаки недостаточности.
23.Глюкозо-аланиновый цикл, его значение при физической работе и голодании.
25.Особенности метаболизма глюкозы в печени, мозге, скелетных мышцах, жировой ткани, эритроцитах.
27.Наследственная энзимопатия глюкозо-6-фосфат-дегидрогеназы. Факторы, провоцирующие проявление недостаточности фермента. Последствия.
29.Роль галактозы в организме. Метаболизм галактозы. Ключевые ферменты метаболизма галактозы. Галактоземия, молекулярные причины, клинические проявления и основы лечения.
31.Гормональная регуляция обмена углеводов. Влияние инсулина, адреналина, глюкагона, кортизола на уровень глюкозы крови и на внутриклеточные процессы превращения глюкозы. Инсулин-зависимые ткани. Гормон- чувствительные ферменты обмена углеводов, способы их регуляции.
33.Общая характеристика сахарного диабета 1 и 2 типа. Нарушенные пути обмена углеводов. Биохимические механизмы осложнений сахарного диабета.