Немного про CRISPR-Cas системы III типа
Мне попались две достаточно интересные статьи про CRISPR-Cas системы III типа. Правильнее сказать, относительно интересные, или интересные для людей с соответствующими интересами. Словом, не всем эта тема покажется интересной, или покажется, но особого отклика не вызовет.
Если попробовать коротко рассказать про этот тип систем, то получится примерно так:
CRISPR-Cas системы III типа - это еще один подвид адаптивных иммунных систем прокариот, которые используют комплементарные взаимодействия между нуклеиновыми кислотами для распознавания и расщепления чужеродных нуклеиновых кислот. В случае систем III типа долгое время было непонятно, что они узнают и как работают, но на текущий момент о механизмах работы этих систем сложилась следующая картина:
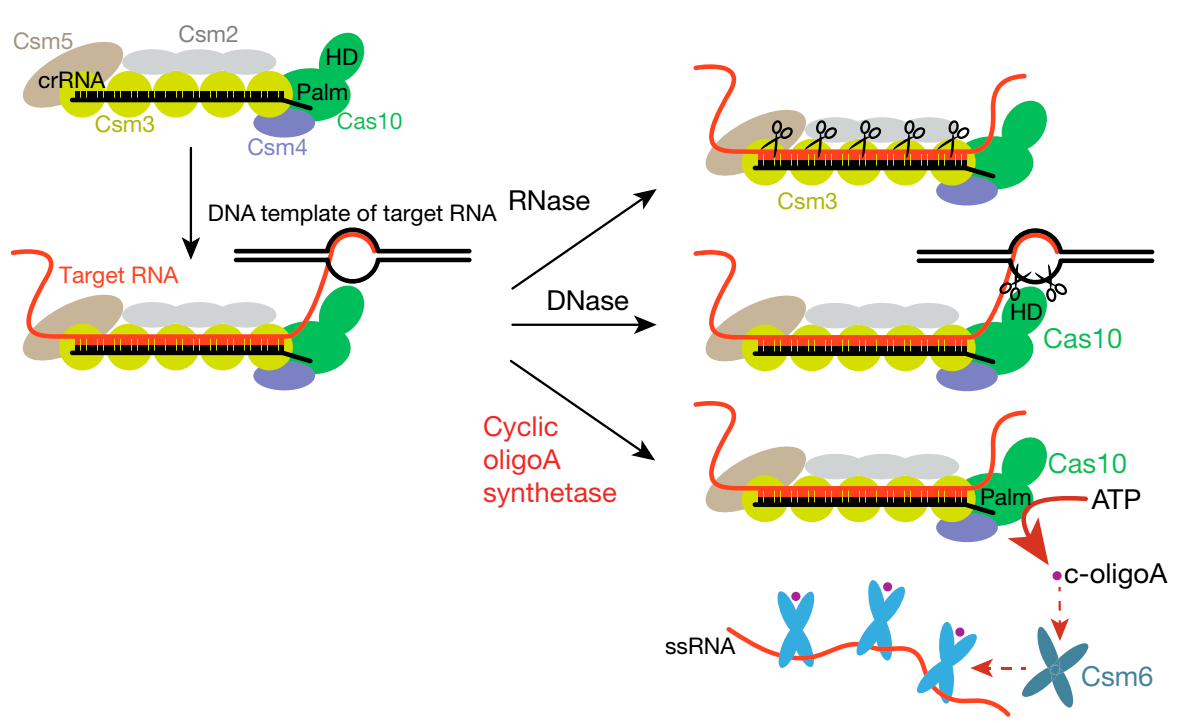
- Cas-гены экспрессируются, давая Cas-белки, CRISPR-кассеты транскрибируются и транскрипты нарезаются на короткие РНК (CRISPR-РНК). Потом несколько Cas-белков связывают эти CRISPR-РНК и образуют эффекторные комплексы. В состав этих комплексов входит большой белок Cas10, который интересен тем, что содержит Palm-домен (как ДНК-полимеразы). Помимо Palm-домена в Cas10 есть еще HD (гистидин-аспартатный типа) домен, который может расщеплять ДНК. Белки, образующие комплекс с CRISPR-РНК (Csm3 на картинке снизу) обладают РНКазной активностью.
- Эффекторные комплексы систем III типа могут распознавать и связывать молекулы РНК, комплементарные CRISPR-РНК; ДНК они не связывают. Когда эффекторный комплекс специфически связывает РНК, это активирует белок Cas10: активированный HD-домен расщепляет одноцепочечную ДНК (предполагается, что это все происходит котранскрипционно, и Cas10 расщепляет ДНК в районе транскрипционного пузыря), а Palm-домен катализирует образование достаточно странных вторичных месседжеров - циклических олигоаденилатов (cOA3, cOA4, cOA6, то есть по 3, 4 и 6 аденилата в цикле). Для этого еще требуется отсутствие тэг-антитэг-комплементарных взаимодействий, но черт с ними, слишком длинно получается.
- Помимо нуклеаз, входящих в эффекторный комплекс, в системах III типа есть еще дополнительные нуклеазы Csm6/Csx1. Эти нуклеазы содержат РНКазный HEPN-домен и так называемый CARF (CRISPR-associated Rossmann fold) домен. CARF-домены связывают циклические олигоаденилаты, и это приводит к димеризации Csm6/Csx1 белков и активации HEPN-доменов (эти HEPN-домены активны только в форме димеров). Активированные нуклеазы Csm6/Csx1 начинают неспецифически расщеплять все РНК в клетке.
- Когда эффекторный комплекс расщепляет распознанную РНК, белок Cas10 выключается - HD-домен перестает расщеплять ДНК, а Palm-домен перестает синтезировать циклические олигоаденилаты. Судьба циклических олигоаденилатов в разных организмах различна - у кого-то есть специальные белки, которые его расщепляют, у кого-то этим занимаются непосредственно нуклеазы Csm6/Csx1 (типа там в CARF-домене какой-то дополнительный нуклеазный центр есть).
Точная роль каждого из этих процессов в иммунном ответе не очень понятна, но по результатам одной статьи (Марраффини) выходит, что ДНКазная активность Cas10 это необходимый компонент (как раз за счет нее система мочит фагов/плазмид), а РНКазная активность Csm6/Csx1 это скорее вспомогательный элемент, который тормозит процессы синтеза компонентов репликации и репарации чужеродных ДНК.
Ну так вот, про статьи, которые мне недавно попались. Первая статья (https://www.biorxiv.org/content/10.1101/784280v1) - про еще одну дополнительную cOA-активируемую нуклеазу, входящую в состав CRISPR-Cas систем III типа в Thermus thermophilus. Только на этот раз это не РНКаза, а ДНКаза, которая может вносить одноцепочечные разрывы в ДНК (специфичность этого процесса пока непонятна). Предполагают, что такие разрывы не очень страшны для бактериальной хромосомы, которая реплицируется относительно медленно, а вот для фагов и плазмид это проблема. Для меня, конечно, эта статья интересна тем, что я сам какое-то время работал с термусами и их фагами, прикольно так.
Вторая статья (https://www.biorxiv.org/content/10.1101/778746v1) - об открытии нового анти-CRISPR белка, который подавляет работу некоторых CRISPR-Cas систем III типа так, что они становятся неактивными против вирусов, которые кодируют такой анти-CRISPR белок. Этот белок открыли в неком вирусе, который заражает архей S. islandicus, и он представляет собой нуклеазу, которая с очень высокой эффективностью расщепляет циклический тетрааденилат, что блокирует активацию Csx1. (Пишут, кстати, что разные Csm6/Csx1 обладают разной специфичностью к разным циклическим олигоаденилатам - одни активируются cOA4, другие - cOA6). Это реально прикольно, поскольку до этого не было известно никаких анти-CRISPR белков против систем III типа. Еще один занятный момент - они пишут, что использовали в работе некий вирус SSeV - "a lytic virus isolated from Kamchatka Russia, (...) (Rowland et al. in prep)". Мне очень интересно, у кого они взяли этот вирус. Все-таки людей, которые изучают термофильных микробов из Камчатки не так уж много, а людей, которые выделяют камчатские термофильные вирусы и того меньше.
Вот такая вот история.