Выбор освещения Ч.3
Внимание! Данный материал содержит информацию, написанную жителем Канады для русскоязычных жителей Канады. Данные научные сведения предназначены исключительно для информационных целей и не являются призывом или пропагандой к употреблению наркотических веществ или к нарушению действующего законодательства.
Пожалуйста, читайте данный материал только если культивация Cannabis Sativa не нарушает законодательство государства, на территории которого вы находитесь в данный момент.
Оглавление
Эффект Красного Обрыва и Эффект Эммерсона
СПЕКТРАЛЬНАЯ ФОТОБИОЛОГИЯ
График эффективности ФАР
Начнем с того, что графики поглощения фотосинтетических пигментов обладают очень низкой репрезентативностью. Предыдущий график фотосинтетических пигментов был нужен лишь для визуализации материала и к реальному спектру работы фотосинтеза отношение имеет весьма условное.
Дело тут в том, что исследования поглощения производятся на пигментах, в большинстве случаев находящихся в таких растворителях, как ацетон или спирт. Совместно с флюоресценцией в реальности мы получаем намного более размытую картину эффективности разных световых диапазонов.
В реальности поглощение листа выглядит как-то так:

Как видно выше, растения эффективно поглощают практически весь свет, попадающий на них, вплоть до отметки в 700 нм.
Оригинальный график МакКри (McCree, 1972) был составлен на основании усредненного результата исследования 20 разных видов растений. Поток света был относительно низким, и точки замерялись отдельными узкими монохромными участками по 25 нм.

У человека может сложиться впечаление, что некоторые фотоны обладают низкой эффективностью, и из-за этого весьма бесполезны. Однако главное, что нужно понять про ФАР карты и все графики относительной эффективности - это закон Энштейна-Старка.
Закон Энштейна - Старка гласит: 1 фотон активирует один электрон. Это означает, что все поглощенные фотоны отдают растению одинаковое количество электронов, что в итоге дает одинаковое количество глюкозы. Разница тут лишь в том, что некоторые спектры поглощаются немного эффективнее, чем другие.
Любой фотон, пойманный антенным комплексом фотосистемы, будет конвертирован в один фотосинтетический электрон через специальную пару хлорофиллов-а.
Исследования, использующие современные способы тестирования, включающие более долгие промежутки освещения и более интенсивные потоки света, приходят к выводу о низкой репрезентативности графика МакКри, о чем будет написано далее.
Диссонанс эффективности синего света
Согласно графикам поглощения отдельных фотосинтетических пигментов, синий свет поглощается намного больше, чем остальные спектры. Согласно же ФАР картам, синий свет используется растениями менее эффективно, чем красный. В чем же причина такой разницы между отдельными пигментами и листом?
В процессе своей эволюции, растения сформировали систему защиты от чрезмерного света. Поэтому такие фотосинтетические пигменты как ксантофилл и каротин компенсируют и рассеивают некоторое количество света, не передавая его специальной паре хлорофиллов-а. Кроме того, не стоит забывать об антоцианине и других флавоноидах, которые в принципе не способны передавать поглощенную энергию для задействования ее в процессе фотосинтеза.
К примеру, в исследовании, при повышении пропорции синего света с 4% до 20% (в 5 раз), урожай линейно падал до -12% относительно контроля. Одной из главных причин было то, что хлорофилл просто получал немного меньшее количество света из-за фотосинтетически-непродуктивного поглощения этой части спектра другими пигментами.
Зеленый свет и его активность
Зеленый свет используется растениями точно также, как синий или красный. Основное недопонимание "низкой эффективности" происходит из-за того, что зеленый почти не поглощается фотосинтетически-активными пигментами. Однако, вместе с этим зеленый свет получает и несколько преимуществ.
В очередной раз сталкиваемся с таким понятием, как флюоресценция. Как видно на изображении ниже, зеленый глубже проникает в лист, так как отражается от хлоропластов немного сильнее, чем остальные спектры.

Чем ярче лампа, тем зеленый свет эффективнее. Происходит это из-за того, что верхние ткани насыщаются синим и красным спектрами, что позволяет зеленому проникать глубже и более эффективно использоваться в процессе фотосинтеза в нижних ярусах листвы.
Эффект Красного Обрыва и Эффект Эммерсона
Эффект Красного Обрыва
Основной проблемой графика МакКри является феномен эффекта красного обрыва. Упрощая, это феномен падения темпов фотосинтетической активности после 680 нм. Его открыл Роберт Эммерсон при тестировании фотосинтетической активности узких промежутков ФАР. Он заметил, что растения практически не используют свет выше 580 нм, когда на них светят узким диапазоном света.

Эффект Эммерсона
Данный феномен как бы является решением проблемы Эффекта Красного Обрыва.
Эффект Эммерсона подразумевает, что для фотосинтеза растения используют красный + дальний красный спектр от 680 до 750нм в том случае, если этот спектр поставляется растениям совместно с более коротковолновым ФАР спектром 400-680 нм.
Происходит этот феномен из-за того, что свет после 680нм активирует исключительно фотосистему I. Проблема же состоит в том, что для фотосинтеза необходима активация и работа обеих фотосистем. Таким образом, фотосистема I не может функционировать без фотосистемы II. Именно поэтому при спектре ниже 680нм работают обе фотосистемы, а после 680нм работает только I.
Последние исследования показывают, что дальний красный свет используется для фотосинтеза точно также, как и остальные участки между 400 и 700 нм. Дальний красный (ДК) имеет эффективность, аналогичную стандартному ФАР диапазону, вплоть до пропорции в 30% ДК от общего диапазона освещения. Далее его эффективность снижается.
Повышенная эффективность дальнего красного света происходит, если к стандартным 1000 PPFD ФАР добавить 300 PPFD дальнего красного, но общий поток посчитать по стандартному диапазону (400-700 нм), что видно по графику (а) ниже. Замеры PPFD будут показывать 1000 PPFD, а темпы роста будут как при 1300 PPFD.

Однако, на графике (b) видно, что если вместо стандартного ФАР диапазона мы используем удлиненный расширенный ФАР (рФАР) от 400 до 750нм, то обьем фотосинтеза увеличивается пропорционально свету идентичного объема в ограниченом диапазоне ФАР.
Оптимальный спектр супплиментраного дальнего красного находится между 730 и 750 нм. В данный момент исследователи пришли к мнению, что процент ДК в общем спектре света для некоторых культур может составлять до 20%, без негативных последствий на общую урожайность растения.
Вывод
Чудес не бывает: давая растению 1000 мкмоль фотонов, мы не можем заставить его реагировать также, как на поток в 1200 или 1300 мкмоль.
Именно по всем вышеперечисленным причинам мы постепенно переходим на новый график фотосинтетической активности, включающий в себя все необходимые исправления. Он хорошо подходит для измерения фотосинтетической радиации, передаваемой в новом диапазоне 400-750нм ePAR (или рФАР по-русски).

ФОТОМОРФОГЕНЕЗ
Фотоморфогенез - это общее морфологическое (структурное) изменение растения под воздействием света, происходящее для адаптации под окружающую среду.
Под понятие фотоморфогенеза подпададает достаточно крупный перечень реакций. Упрощая, их все можно разделить на два основных типа:
Синие реакции
Высокая энергия синего света вызывает общий каскад реакций компенсации высокой освещенности. Именно поэтому, при активации фоторецепторов синей реакции мы видим, что обилие света в диапазоне между 300 и 500нм вызывает редукционный фотоморфогенез (иными словами - сжатие).
Последствия редукционного фотоморфогенеза включают такие процессы, как:
Синие реакции стимулируют выработку таких фотопротекционных молекул, как флавоноиды, концентрацию которых часто невооруженным глазом видно из-за обилия антоцианинов.
Данный феномен гипотетически весьма просто объясняется тем, что растение, подвергаясь высоким уровням синего света, пытается оптимизировать метаболизм, не подвергая себя чрезмерному стрессу. Чем больше слоев оно нарастит, тем меньше урона оно получит и тем эффективнее сможет усваивать попадающий на листву свет.
Красные реакции
Энергия красного света вызывает реакции компенсации низкой освещенности. Суммарный эффект от активации красных рецепторов в диапазоне между 500 и 780нм вызывает эффект избегания тени.
Эффект избегания тени включает:
- Расширение и утоньшение листвы,
- Удлинение стеблей,
- Поднятие листьев для улучшенного улавливания света,
- Ускорение переходов между вегетацией и цветением.
Данный феномен гипотетически объясняется тем, что растение, в условиях красного спектра, пытается компенсировать недостаточное количество энергии, а также перегнать конкурирующих соседей, которые отбрасывают на него тень.
Фоторецепторы
В отличие от фотосинтетических пигментов, которые расположены в хлоропластах, фоторецепторы чаще всего представляют из себя свободно плавающие в разных частях клетки белковые структуры.
Они практически являются глазами растений. Благодаря им каннабис может ощущать и "видеть" пространство вокруг себя, определять количества получаемого света, а также замерять длину дня.
Кроме базовых синих и красных реакций, каждая группа фоторецепторов имеет множество уникальных для своего типа функций. На данный момент известно 4 основные группы фоторецепторов:
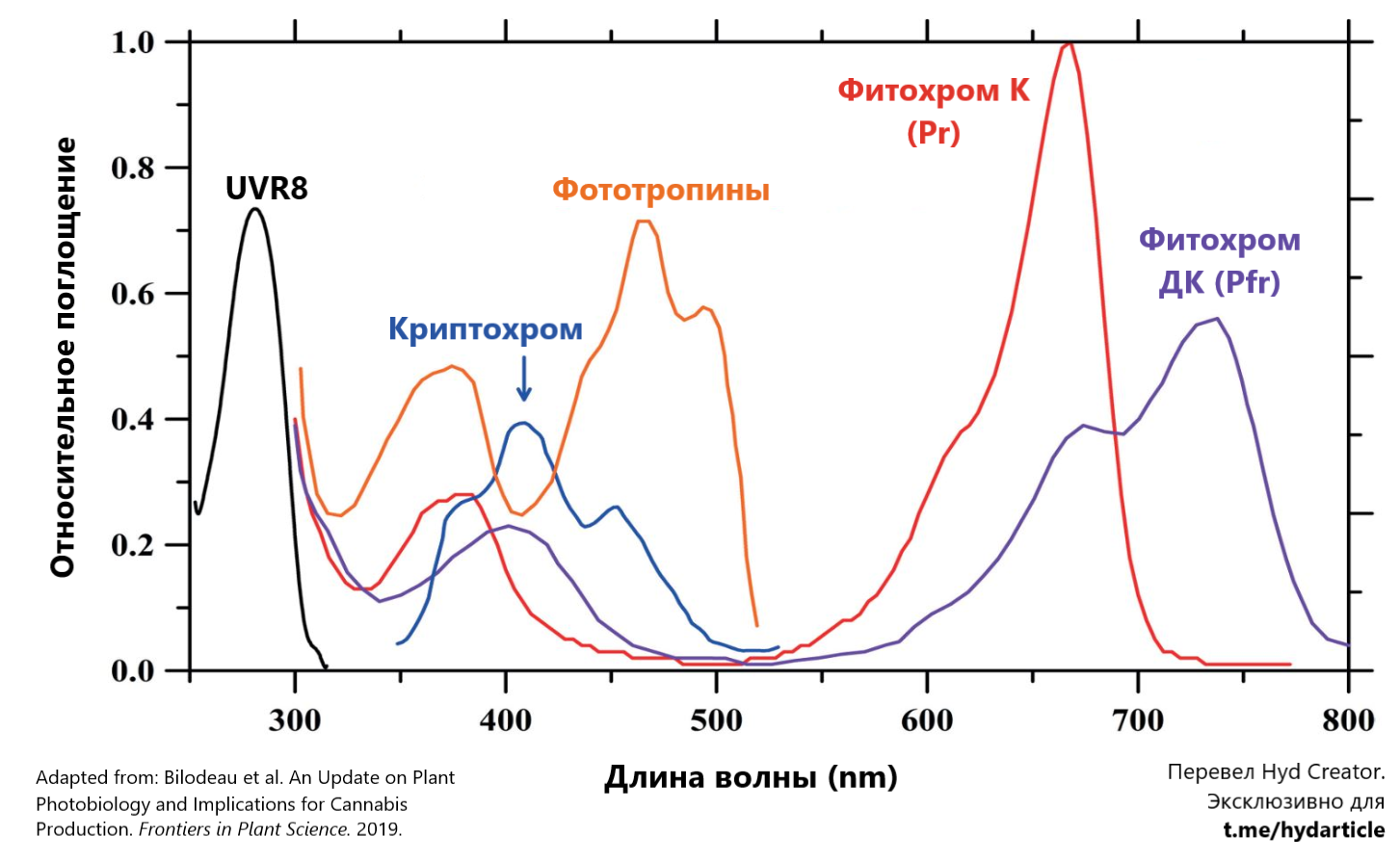
UVR8 (UV Resistance locus 8)
Пик поглощения ~290нм. Синяя реакция.
Восьмой локус резистентности ультрафиолету (UVR8) - это фоторецептор, который был открыт в качестве главного маркера, реагирующего на ультрафиолет диапазона B. Однако, позже исследователи выяснили, что не смотря на то, что UVR8 является главным индикатором получения UV-B радиации, он также может реагировать на многие другие условия среды, включая: UV-A, температурный стресс, засоленность субстрата, инфекции и интоксикацию тяжелыми металлами.
Таким образом, этот фоторецептор сигнализирует растению о широком перечне экстремальных ситуаций. Делает он это для контроля починки ДНК. Исследователи проходят к выводу, что активацию UVR8 растения распознают как чрезвычайный сигнал, который заставляет приостанавливать многие жизненные процессы и бросать все силы починку ДНК.
Касательно эффекта УФ-Б на растения в общем, есть результаты исследований, показывающих, что данный диапазон ЭМ излучения может иметь длинный список негативных воздействий на растения. В эффекты UV-B радиации входят: запуск механизмов починки ДНК, разрушение фотосистем I и II, снижение активности РУБИСКО, урон тилакоидным мембранам хлоропласта, разрушение хлорофилла и каротеноидов, сокращение биомассы, а также снижение урожайности.
В недавнем эксперименте на каннабисе ученые выяснили, что низкие уровни УФ-Б практически не влияли на итоговое состояние растений, а постоянное освещение каннабиса высоким количеством ультрафиолета линейно снижало как урожайность, так и содержание каннабиоидов в соцветиях.

Криптохром (Cryptochrome)
Пик поглощения ~410нм. Синяя реакция.
Основная задача криптохрома - это помогать кроне фотосинтезирующиего организма измерять и понимать примерные количества света, которые получает растение.
Регулирует циркадные ритмы растения, включая процессы фототропизма и фотопериодизма. Управляет транспирацией за счет контроля раскрытия стоматы (устьиц). Блокирует реакцию растения на активацию фитохрома, не давая происходить красной реакции (избегания тени) при высоких уровнях синего света.
Фототропины (Phototropins)
Пик поглощения ~460нм. Синяя реакция.
Как следует из названия, фототропины регулируют процесс фототропизма и, в том числе, фоторелокацию. Они отвечают за регуляцию циркадных ритмов растения и, также как и криптохром, контролируют раскрытие стоматы (устьиц).

Как видно из изображения выше, отрицательный фототропизм корней контролируется синим и зеленым спектрами и не реагирует на красный. Это указывает на важность спектра активации фототропинов в регуляции фототропизма.
Фитохромы (Phytochrome)
Пики поглощения ~660+735нм. Красная реакция.
Фитохром нужен растению для регуляции фотопериодизма и процесса измерения освещенности. Существует две разных формы фитохрома, которые постоянно переходят одна в другую:
Фитохром, поглощающий красный свет (К). Неактивная форма, имеющая пик поглощения около 660нм.
Phytochrome far red (Pfr или Фдк)
Фитохром, реагирующий на дальний красный свет (ДК). Главный активный маркер, который используется растением для контроля фотоморфогенеза и фотопериодизма. Имеет пик поглощения около 740нм.
Под воздействием красного спектра, красный Фк переходит в форму Фдк. Под воздействием дальнего красного Фдк переходит в форму Фк.

На графике выше видно соотношение форм фитохрома и Красного/Дальнего Красного. Чем больше короткого красного света от всего красного диапазона (К+ДК), тем больше формы Фдк от общего количества фитохрома.
Как видно из графика, светодиодные панели даже со специальным добавлением дальнего красного дают намного более высокое соотношение неактивного Красного, чем, к примеру, прямой солнечный свет или солнечный свет в теплице.
Дальний красный фитохром намного менее стабилен, чем базовый красный, и поэтому он распадается ночью. По его концентрации растение замеряет длину ночи, что контролирует процесс фотопериодизма.
Переход фитохрома между формами блокируется высокой освещенностью, и тем самым он застревает в промежуточной форме. Это вызывает скачок концентрации Фдк в растении после выключения света. Ощущение большой разницы между начальным количеством фитохрома и конечным (утренним) позволяет растению отслеживать продолжительность дня.
Манипулирование спектром
В коммьюнити сформировалось общее понимание такой теории, как спектральная манипуляция. После прочтения части про фоторецепторы у гровера может сложиться впечатление, что манипулировать морфологией и химическим составом растения не составляет никакой трудности.
Давайте разберемся, насколько эта мысль совпадает с реальностью.
Стимуляция ультрафиолетом
Влияние UV-B диапазона на морфологию и урожайность каннабиса уже было частично проанализировано в части об UVR8. Так как же все-таки ультрафиолет влияет на химический состав каннабиса?
Думаю первое, на что стоит обратить внимание - это отсутствие подтверждений гипотезы о том, что А и Б ультрафиолет увеличивает содержание ТГК в соцветиях каннабиса.
Основное исследование, которое используется в качестве доказательной базы в 99% случаев, было проведено в 1987г на низко-каннабиноидных сортах (1-3% ТГК) с использованием UV-B. Хоть оно и показало среднее увеличение каннабиноидов с 2.5 до 3.2%, сами авторы заявили о неоднозначности полученных результатов.

Второе, более современное исследование 2018 года показало небольшое увеличение содержания каннабиноидов с использованием ультрафиолета, но проблема в том, что в нем сравнивались слабые количества света (450 PPFD).
Третье исследование пришло к выводу, что супплиментарное освещение УФ-А и УФ-Б не увеличивает содержание ТГК в соцветиях, однако вызывает увеличение трихом и каннабиноидов на 30% в сахарных листочках. Весьма интригующие результаты, однако вновь проблема может заключаться в том, что исследование проводилось на относительно невысоких уровнях света (600 PPFD). Положительных результатов большей выработки ТГК от применения УФ-А на соцветиях каннабиса обнаружено не было.
Возможной причиной отсутствия реакции на ультрафиолет является феномен достижения биологических лимитов растения. Генетический потенциал синтеза каннабиноидов у многих современных сортов с >25% ТГК находится очень далеко от среднего содержания ТГК в лендрейсах. Тем самым он подходит к "пределу" своих возможностей с каждым выведеным сортом.
К примеру, в том же самом исследовании из 1987, сами авторы предполагают, что концентрации КБД в 2.3% в тканях технического каннабиса было достаточно для защиты от ультрафиолетового излучения диапазона UV-B.

В научной работе 2021 года исследователи сформулировали гипотезу, что ультрафиолет может являться условием, стимулирующими долгосрочную адаптацию растений.
К примеру, сорта из высокогорных регионов Афганистана, подвергнутые повышенному количеству ультрафиолета, сотни лет адаптировались под жёсткие условия среды. Именно поэтому они стали более низкими, быстроцветущими и смолистыми, нежели чем их родственники из тропических регионов. Они попросту имели тысячу предыдущих поколений, которые постепенно меняли свое ДНК для более успешного размножения в суровом ареале своего обитания.
Как видно из графика спектрального поглощения, все фоторецепторы, кроме криптохрома, обладают вторичными пиками в диапазоне 300-400нм. Это позволяет свету в УФ-А спектре вызывать такую же синюю реакцию в растении, как и стандартный длинноволновый синий свет. Иными словами, УФ излучение стимулирует синтез флавоноидов.
В том же исследовании из 2021, ученые пришли к выводу, что освещение каннабиса УФ-Б диапазоном линейно снижало содержание терпенов, также как урожайность и содержание каннабиноидов.
Что касается исследований других растений, в них прослеживается тенденция на общее изменение терпенового профиля. В то время как концентрации некоторых терпенов растут, другие могут значительно снижаться.
Однако, нам необходимо больше независимых исследований для того, чтобы делать какие-то выводы. Вполне возможно, что влияние УФ-Б на терпеновый профиль контролируется генетикой. Это означает, что подобная стимуляция, в зависимости от сорта, может как приводить к увеличению общего количества терпенов, так и уменьшать их концентрацию.
Соотношение красного и дальнего красного
Уже несколько раз в этой части статьи упоминался эффект дальнего красного спектра на морфологию растений. Так как же и почему происходит эффект избегания тени?
Основная причина такой уникальной реакции на дальний конец фотосинтетически-активного спектра заключается в особенном взаимодействии этого диапазона с клетками растения. В то время как красный свет эффективно поглощается антенными комплексами хлоропласта, дальний красный практически не поглощается пигментами и, следовательно, отражается гораздо сильнее.
Когда растение получает относительно высокие количества дальнего красного, это деактивирует дальний красный фитохром, переводя его в красное состояние. Чем больше дальнего красного света попадает на растение, тем меньше дальнего красного фитохрома остается в клетках. В итоге, растению начинает казаться, что оно находится в тени других растений, либо окружено большим количеством конкурентов.
Побег старается всеми силами перерасти своих соседей, чтобы получить больше света. То же самое происходит и с переходами между жизненными этапами: преждевременное зацветание гарантирует меньшую конкуренцию за свет, воду, ресурсы и пыльцу.

Часто цитируемые исследования на салате показывают, что замена всего 15% обычного ФАР потока на поток в 700-750нм увеличиает урожай на 30%. Проблема тут в том, что урожайность салата образуют его вегетативные ткани (листва). Увеличение урожайности происходит из-за увеличения площади листвы, что в последствии увеличивает количества пойманого света.
Однако, чуда произойти не могло даже и с салатом. С увеличением урожайности количество хлорофилла в листьях упало на 20%, что вызвало снижение эффективности улавливания фотонов на 14%.
Взаимодействие красной и синей реакции
На сегодняшний день существует множество опытов, согласно которым растения иначе реагируют на различные спектры.
Современные исследования (1 и 2) приходят к выводу, что манипуляция ФАР спектром для воздействия на морфологию или стимуляцию синтеза каннабиноидов работает исключительно при низких обьемах света. Что касается средне-высоких уровней PPFD, гипотеза состоит в том, что такой способ манипуляции теряет эффективность из-за того, что фоторецепторы насыщаются.
По части морфологии, фоторецепторы синей реакции сигнализируют растению о высокой освещенности, фоторецепторы красной - о слабом уровне света.
Чем больше света, тем слабее растение склоняется к красной, либо синей реакции. Таким образом, синие и красные рецепторы соединяют свои показания, блокируя излишние ответы друг-друга, тем самым давая растению равномерно развиваться и понимать, что оно получает достаточные количества света.
Подводя итог в вопросе спектров
Ближний ультрафиолет, которым обладают большинство лед ламп, все равно находится слижком близко к концу синего света и попадает под поглощение как фотосинтетическими пигментами, так и синими фоторецепторами.
Иными словами, ультрафиолет вплоть до 350нм просто используется в процессе фотосинтеза и помогает растению лучше понимать количество получаемого света, и, следовательно, лишь запускает синюю реакцию, тем самым увеличивая количество флавоноидов. УФ-Б имеет потенциал к увеличению эфирных масел и терпенов, однако необходимо больше тестов.
Доказательств того, что оба этих спектра способны увеличивать концентрации ТГК, у нас нет.
Целесообразность супплиментации ДК спектром очень зависит от культивируемого вида растений. Не каждый гровер захочет видеть увеличенное нарастание листвы и удлинение стеблей в ограниченных пространствах бокса или гроу-рума. Кроме того, уже знаменитые исследования на салате, гласящие об увеличении урожайности на 30%, производились при уровнях света в 350 PPFD, что снова наводит на мысль о том, что реакция растений на спектр может нивелироваться восокими объемами фотосинтетически-активной радиации.
Не стоит также забывать, что ваши растения получают дальний красный не только от специальных диодов. Обычный белый спектр с цветовой температурой в 3500К добавляет около 3% ДК света. Вместе с тем, каждый отдельный куст в вашем руме все время будет получать умеренные сигналы затенения от своих же соседий.
Для получения большей выгоды эффективнее всего давать растению наибольший объем сбалансированного спектра (условные 4000К). ДК и УФ-А светодиоды являются приятным бонусом, но специально докупать панели, которые дают дополнительный УФ-А, либо каким-то специальным образом "стимулируют цветение", смысла не вижу (по крайней мере, в малых масштабах).
Подобные "стимуляторы" и "усилители" всего лишь дают растению немного больше спектра в диапазонах 660 и (иногда) 750нм. Эти спектры никак самому цветению не помогают, а просто увеличивают общий объем фотосинтетики, как и любой другой свет в диапазоне 350-750нм. Кроме того, такие "бустеры" всегда дороже простого "белого" борда аналогичной мощности.
Красный используется в процессе фотосинтеза, но никак не стимулирует цветение!
Даже больше, на данный момент можно найти мнения крупных производителей, которые говорят о том, что высокое количество красного света снижает урожайность каннабиса. Резюмируя, использование бустеров может даже вредить.
ФОТОТРОПИЗМ
Фототропизм - это тип фотоморфогенеза, определяющий движение растения относительно падающего на его ткани света.
Разделяется на 2 главных типа: положительный и отрицательный.
- Положительный фототропизм подразумевает движение растения к свету. (Выравнивание листьев в зависимости от угла падения света).
- Отрицательный фототропизм - движение растения от света. Это ключевой механизм ориентации для корней, растущих в сторону наименьшей освещенности.
В стандартных условиях корневая зона развивается еще и благодаря гравитропизму (движения к гравитации), однако именно отрицательный фототропизм помогает растениям развивать корневую систему в условиях невесомости.

Сам механизм удлинения происходит засчет активного транспорта ауксинов (фитогормонов) в ткани, находящиеся ниже точки роста для стеблей и выше точки роста для корней. Таким образом происходит удлинение клеток, которое и вызывает сгибание или поворот растущей части к свету, либо от неё.
ФОТОПЕРИОДИЗМ
Фотопериодизм - это способность растения подстраивать свои биоритмы под протяженность светового дня. Растение не измеряет количество часов света, ему важна длина ночи, а именно - сигналы от фитохрома, криптохрома и фототропинов.
Наступление темноты сигнализирует растению о завершении дня и запускает переход накопленного за день Pfr в Pr. Когда количества Pfr уменьшаются на постоянной основе, растение понимает, что темного времени стало больше и запускает цветение.
Именно поэтому старой хитростью для перевода затормозившего растения является установка 36ч непрерываемой темноты. Таким образом, максимально возможное количество Pfr разрушится до состояния Pr, что послужит четким сигналом о смене сезонов.
Криптохром и фототропины тоже помогают растениям измерять протяженность дня. У нас нет данных, что исключительно красный свет запускает переход между фазами. Как красная, так и синяя части рФАР важны для процесса фотопериодизма.
В современном гроверском коммьюнити устоялись стандартные диапазоны освещения каннабиса в 18 часов на вегетации и 12 часов света на цветении.
Однако, как мы знаем, разные места на поверхности нашей планеты, в зависимости от широты и сезона, порой имеют весьма далекую от 18 и 12 протяженность светового дня. Тем не менее, дикий каннабис стабильно растет и развивается в этих регионах.
Следовательно, растения, формировавшиеся в разных широтах, переводятся на цветение разными длинами светового дня. Тут в игру вступает понятие критического фотопериода. Простыми словами, критический фотопериод - это суммарное количество часов темноты, при котором растение начинает зацветать.

Зная критический фотопериод сорта и длину светового дня на нашей широте, мы можем предсказать, будут ли выбранные нами растения успевать развиваться так, как нам хочется.
- Сорта со слишком коротким критическим фотопериодом могут не успевать зацветать в нашей широте до наступления осенних заморозков.
- Растения со слишком длинным критическим фотопериодом, возможно, не будут иметь достаточно времени для вегетации и моментально зацветут, что поставит урожайность под удар.
Существует тенденция, что сорта, генетика которых основана на более дальних от экватора лендрейсах, в среднем имеют более короткий критический фотопериод. Сорта же из более близких к экватору регионов зацветают при больших длинах ночи.
Как видно из графика ниже, стандарный критический фотопериод для фотопериодных растений в среднем находится в диапазоне между 13 и 15 часами света.

Зная критический фотопериод культивируемого сорта гровер может добавить некоторое количество света в световой день. В теории, простое увеличение фотопериода на цветении способно добавить от 8 (при 13 часах света) до 25 (при 15 часах света) процентов DLI в день при одинаковом количестве PPFD.
Касательно фотопериода во время вегетативного периода, все зависит от осветительного оборудования и DLI, о котором речь пойдет дальше. При достаточной мощности света гровер может использовать как стандартные 18/6 для экономии электроэнергии, так и компенсировать недостаточное освещение с помощью удлинения фотопериода до 20 часов.
Для экспериментов со временем светового дня сначала нужно понять, что при более длинном световом фотопериоде растения имеют склонность дольше зацветать, и вместо стандартных 5-7 дней для проявления первых признаков могут потребовать от 10 до 15 дней. Также не стоит забывать, что световой день, находящийся на грани критического фотопериода, может сделать цветение нестабильным, что поставит под удар качество урожая.
Если не хотите рисковать, то попробуйте переводить растения с 20/4 на 13/11 часов. Для большинства сортов этого сигнала должно хватить для четкого перехода и стабильного цветения.
Для проведения более рискованных экспериментов необходимо иметь идеальные растения без каких-либо симптомов стресса. Для нахождения критического фотопериода своего сорта, попробуйте вместо перевода на привычной неделе №6 перевести растение с 20 до 14 часов света на неделе №4. Подождите 10 дней, и если изменений не последовало, то устанавливайте стандартные 12 часов.
Дальний красный на ночь
Последнее время становится популярным освещать растения дальним красным спектром сразу после отключения света. Когда в естественных условиях солнечные лучи преломляются, растение получает высокую дозу дальнего красного, что мгновенно переводит некоторое количество дальнего красного фитохрома в неактивную (красную) форму. Таким образом гроверы пытаются копировать естественный закат и давать толчок естественному разрушению ДК фитохрома.
Доказательств эффективности использования этой технике на каннабисе в данный момент нет. Исследования скорее указывают на то, что подобная практика работает только на растениях длинного дня, зацветающих при увеличении светового дня (весной).
Кроме всего прочего, высокая освещенность меняет механизм конвертации фитохрома, так что все опять может упираться в общую мощность светового потока. Более сильный свет, в теории, может блокировать больше фитохрома днем, что будет давать больший его скачок после наступления ночи.
Данный инструмент бесполезен, если вы используете стандартные фотопериоды (18/6 + 12/12), либо просто выращиваете несколько разных сортов одновременно.
В перспективе это может послужить способом тонкой настройки критического фотопериода, если вы хотите давать растению максимально длинный день. Однако пока что данная гипотеза не представляет большей ценности, чем простое предположение.
Свет и потенциал вреда
Теперь стоит поговорить о двух вызываемых светом процессах, имеющих потенциал снижения фотосинтетической эффективности.
Фотоингибиция
Фотоингибиция - это неизбежный процесс, происходящий в любом растении во время активного фотосинтеза. Упрощая, фотоингибиция переводит хлоропласты в восстановительную фазу путем остановки процесса фотосинтеза. В большей степени ему подвержена именно Фотосистема II, так как она занята самой интенсивной частью световой фазы фотосинтеза (разламыванием воды).
Так как любой инструмент подвержен постепенной деградации в условиях высокой нагрузки, темпы фотоингибиции прямо пропорциональны уровням фотосинтеза. Когда света много, кроме потребности в ресурсах для роста растение также начинает нуждаться в повышенных средствах для починки своих фотосистем.
При стандартном течении метаболизма хлоропласт все время отключает (фотоингибирует) и чинит вышедшие из строя структуры. Однако, если фотопротекционные пигменты (каротин, ксантофилл и антоцианины) перестают защищать фотосистемы от чрезмерного урона, то баланс фотоингибиции и восстановления теряет равновесие.
Таким образом, останавливается фотосинтез и растение начинает тратить запасенную энергию для гашения возбужденных пигментов и починки фотосистем вместо углеродной фиксации.
Основными спектрами, вызывающими фотоингибицию, являются высоко энергетичные синий и ультрафиолетовые диапазоны. Именно поэтому, при использовании богатых этими спектрами источников освещения, мы видим замедление метаболизма и редуктивный фотоморфогенез .
Фотооксидирование
Необратимый процесс разрушения фотосистемы.
Иногда антенному комплексу не удается передать полученную световую энергию дальше. К примеру, если повреждается реакционный центр фотосистемы, то в дело вступает фотоингибиция. Если же фотоингибиции не хватает ресурсов или времени для безопасного высвобождения полученной энергии и своевременной починки фотосистемы, то полученный из разламывания воды кислород начинает окислять близлежащие структуры. Происходит цепная реакция выгорания хлоропластов вместе со всеми содержащимися в них пигментами.
У процесса фотооксидации есть 3 основных причины:
- Неспособность криптохрома и фототропинов адекватно понимать и подстраивать растение под количество получаемых ФАР фотонов. К примеру, когда растение освещается чрезмерным количеством красного света, который на эти синие рецепторы не воздействует.
- Запредельные уровни света. Хоть каннабис и способен выдерживать колоссальные нагрузки в виде 2000 мкмоль света на постоянной основе, переход PPFD за черту в 2100 может вызывать выгорание.
- Генетическая предрасположенность или мутация, влияющая на предыдущие два пункта. Разные сорта имеют разные способности к контролю света.

Качество соцветий, подвергнутых выгоранию, очень зависит от изначальных параметров выращивания, генотипа, хемотипа, а также стадии, на которой произошло выгорание.