Энергия мышц
Для достижения спортивных результатов в соревнованиях, длящихся от секунд до нескольких часов, необходимо непрерывное снабжение АТФ фундаментальных клеточных процессов, лежащих в основе сокращения скелетных мышц во время физической активности. Поскольку запасы АТФ в мышцах невелики, все пути метаболизма должны быть активированы с целью поддержания требуемой скорости ресинтеза АТФ. Эти пути включают расщепление фосфокреатина и мышечного гликогена, что обеспечивает анаэробное фосфорилирование и окислительное фосфорилирование за счет метаболизма углеводов и жиров.
Относительный вклад этих метаболических путей в первую очередь определяется интенсивностью и продолжительностью тренировки или старта.
Для большинства спортсменов углеводы являются основным источником топлива для анаэробного и аэробного метаболизма.
В обзоре, опубликованном в журнале Nature Metabolism, авторы рассматривают ключевые пути метаболизма и пути его регуляции при физической нагрузке, гарантирующие ресинтез АТФ, точно соответствующий потребности.
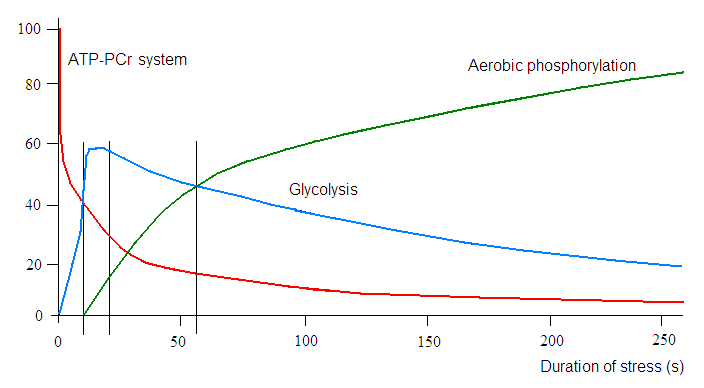
Не секрет, что любые спортивные результаты определяются сложным взаимодействием биологических, психических факторов и факторов окружающей среды. Доступность АТФ имеет решающее значение для сократительной активности скелетных мышц, как в соревнованиях, где нужна взрывная сила или спринт, длящийся секунд или минуты, так и в соревнованиях на выносливость, порой длящихся часами. АТФ необходим для активности ключевых ферментов, участвующих в работе клеточных мембран, обмена кальция внутри клетки и механизма мышечного сокращения. Поскольку внутримышечные запасы АТФ относительно малы (~ 5 ммоль на кг мышечной ткани), они не могут поддерживать сократительную активность в течение длительного времени. Например, во время полной максимальной нагрузки (например, во время спринта) при выходной мощности 900 Вт (~ 300% VO2 max) расчетная скорость использования АТФ составляет 3,7 ммоль АТФ кг/с, и если бы запасенный АТФ был единственным источником энергии, такой спринт мог бы длиться лишь <2 с. Во время субмаксимальной нагрузки ~ 200 Вт (~ 75% VO2 max) соответствующие значения составляют 0,4 ммоль АТФ кг/с и ~ 15 с, соответственно.
Следовательно, должны быть активированы другие метаболические пути, включая анаэробное или аэробное фосфорилирование. Последнее в значительной степени зависит от дыхательной и сердечно-сосудистой систем, гарантирующих адекватную доставку кислорода к сокращающимся скелетным мышцам и от наличия адекватного количества метаболитов, в первую очередь углеводов и жиров. Анаэробные энергетические пути имеют гораздо большую мощность (скорость производства АТФ), но меньшую мощность (общее количество произведенного АТФ), чем аэробные пути. Что касается окислительного метаболизма, окисление углеводов имеет более высокую выходную мощность, но меньшую мощность, чем окисление жиров; это один из факторов, способствующих снижению выходной мощности при истощении углеводов во время длительных нагрузок.
Оба (анаэробный и аэробный) пути активируются одновременно с самого начала активности, чтобы точно соответствовать требованиям каждой ситуации. Хотя аэробные пути являются стандартными, доминирующими по выработке энергии во время тренировок на выносливость, им требуется время (от секунд до минут), чтобы полностью активироваться, в то время как анаэробные системы быстро (от миллисекунд до секунд) обеспечивают организм энергией, покрывая потребности в той ситуации, когда аэробная система не может их обеспечить. Анаэробная энергия важна во время высокоинтенсивных активностей, таких как спринт, когда потребность в энергии намного превышает скорость, которую могут обеспечить аэробные системы. Эта ситуация очень часто встречается в спорте с постоянными остановками, когда переходы от “низкоэнергетических” к более высоким потребностям многочисленны, и в этом случае включение двух систем и обеспечение как аэробной, так и анаэробной энергии способствует достижению успеха. В совокупности, производство аэробной энергии с использованием жиров и углеводов в качестве топлива, а также обеспечение анаэробной энергии за счет распада фосфокреатина и использования углеводов в гликолитическом пути позволяют спортсменам удовлетворять высокие энергетические потребности в различных видах спорта.
Различные метаболические пути регулируются рядом внутримышечных и гормональных сигналов, которые влияют на активацию ферментов и доступность субстратов, тем самым гарантируя, что скорость ресинтеза АТФ близко соответствует потребностям АТФ при физических нагрузках. Регулярные тренировки и различные диетические вмешательства активно используются для повышения устойчивости к усталости за счет изменения доступности субстратов и воздействия конечных продуктов метаболизма.


Одна из проблем в изучении регуляции метаболизма при физической нагрузке в скелетных мышцах заключается в том, что большая часть доступных данных была получена в результате исследований на мужчинах. Хотя основные принципы, контролирующие регуляцию метаболизма, верны как для женщин, так и для мужчин, были отмечены некоторые различия. Работа с хорошо тренированными легкоатлетами не выявила различий в активности ферментов скелетных мышц, составе волокон и окислении жиров между мужчинами и женщинами. Однако, более поздние исследования показали, что больший процент потребления топлива у женщин, тренирующихся с такой же относительной субмаксимальной интенсивностью, приходится на жир, и этот эффект, вероятно, связан с уровнями циркулирующего эстрогена. У женщин более высокая доля волокон типа I, более низкая максимальная емкость гликолитических ферментов и большая зависимость от внутримышечных триглицеридов (IMTG) во время тренировок, чем у мужчин, эффекты, которые могут быть связаны с более высокими концентрациями IMTG у женщин в волокнах первого типа. Кроме того, введение эстрогена мужчинам снижает окисление углеводов и увеличивает окисление жиров во время тренировок на выносливость. Эти результаты показывают, что женщины могут лучше “подходить” к тренировкам на выносливость, чем мужчины.
Еще одна область, которая была исследована, - это влияние фазы менструального цикла и менструального статуса на регуляцию метаболизма скелетных мышц. Как правило, исследования, изучающие тренировки в лютеиновой и фолликулярной фазах, сообщили только о незначительных изменениях или отсутствии изменений в жировом и углеводном обмене при различной интенсивности тренировки.

Понимание энергетического обеспечения тренировок, регуляции метаболизма и использования жиров и углеводов во время тренировок расширилось за более чем 100 лет исследований. Однако практически во всех областях регуляции жирового и углеводного обмена, многое остается неизвестным, а возможность полностью перенести результаты экспериментов in vitro и исследований на животных на тренировки людей остается ограниченной. Остается много вопросов во всех областях энергетического обмена, регулирования жирового и углеводного обмена во время тренировок, оптимальных тренировочных вмешательств и возможности манипулирования метаболическими реакциями для достижения эргогенных преимуществ. Таким образом, биология тренировок будет оставаться плодотворной областью исследований в течение многих лет, поскольку исследователи стремятся лучше понять метаболические основы спортивных успехов.